(5 milionů – 40 000 př.n.l.)
vznik a vývoj člověka
kreacionismus – v
širším smyslu slova přesvědčení,
byly stvořeny zvláštním
zásahem vyšší bytosti či božstvem
termín „striktní
kreacionismus“ se někdy používá
pro nábožensky motivované
odmítání teorie evoluce,
aby nedošlo k mylné záměně se
všeobecnějším chápáním stvoření,
zastávaným věřícími, jejichž
náboženská víra akceptuje teistickou evoluci
podle většiny přírodovědců
je striktní kreacionismus netestovatelná nevědecká
teorie
a jako taková nemůže být předmětem vědeckého zkoumání
ani součástí vědeckých teorií
přírodovědecké práce pracující s aspektem
speciálních nadpřirozených zásahů do evoluce
jsou obvykle považovány za pseudovědecké
zastánci striktního kreacionismu
naopak zpravidla odmítají vědecké závěry
presentující evoluci jakožto nejpravděpodobnější vysvětlení
existence známého universa
na událost samu (tj. cílený zásah) kreacionisté nahlíží
buď jako na akt stvoření ex nihilo, tedy z ničeho,
anebo jako vznik pořádku, jemuž předcházel chaos,
který někdo uspořádal (demiurg)
ačkoli víru lze interpretovat i doslovně (tj. fyzikálními a biologickými
pojmy),
náboženské diskuse se většinou omezují na
duchovní rozměr problému
kreacionismus tedy jednoduše zdůrazňuje
duchovní podstatu lidských bytostí tím,
že ji považuje za základ celé přírody
a označuje všechny ostatní úhly pohledu za materialistické,
ignorující duchovní principy
mnozí lidé věřící ve stvoření
považují toto přesvědčení
za součást náboženské víry
a tudíž je z jejich hlediska mimoběžné s pohledem vědeckým
jiní prosazují názor,
že vědecká pozorování jsou v souladu s ideou stvoření
zastánci tzv. teistické evoluce považují vědecky popsané
mechanismy
za projev stvořitelského díla
věřící orientovaní spíše na vědu
vnímají popis stvoření v náboženských textech
jako oslavný hymnus na celé stvoření,
který nikdy nebyl zamýšlen jako historická výpověď
striktní kreacionismus dnes
přežívá
v některých
fundamentalistických náboženských společenstvích –
u křesťanů např.
v některých konzervativních protestantských denominacích
či sektách,
trvajících tedy na doslovném
výkladu textu Písma – knihy Genese
např. v některých,
zejména jižních státech USA –
jejich snaha
o prosazení vyučování kreacionistického pohledu
vedle evoluční teorie
v rámci
vědeckých disciplín není úspěšná
u nás se
nejnověji deklaroval jako striktní kreacionista
kancléř
presidenta Klause Petr Hájek
(„jako
že je Bůh nade mnou, já z opice nepocházím!“)
kreacionismus
bývá někdy směšován s teorií inteligentního designu –
je
však potřebné obě koncepce odlišovat
kreacionismus
vychází z předpokladu, že svět byl stvořen bohem,
kdežto
teorie inteligentního designu
na
základě analýzy biologických fenoménů
pouze
dospívá k závěru,
že
tyto fenomény v sobě nesou prvky účelnosti a záměru –
tedy
inteligence
na
otázku, kdo je nositelem této inteligence, teorie neodpovídá,
neboť
tato otázka je svou povahou metafyzická
evoluční teorie,
vychází z předpokladu
uplatňování přirozeného výběru v živé přírodě
Charles Darwin
– O původu druhů
darwinismus i evoluční teorie
často byly
a někdy dosud jsou zpochybňovány
z náboženských hledisek
–
zde moderní křesťanství ovšem
rozpor nevidí –
cf. jezuitský filosof a
antropolog Teilhard de Chardin
a jeho podíl na zkoumání procesu
antropogeneze
(výzkumy v jeskyni
Ču-ku-tien u Pekingu)
postupně ale kritika darwinismu
také ze strany některých věd –
genetika, která tvrdila, že
mutace genů jsou nahodilým jevem,
který tudíž nevyvolává žádný
pokrok –
podcenění přirozeného výběru
moderní biologie dokázala
evoluční teorii uvést do souladu
s aktuálním stavem
poznání genetických zákonitostí
evoluce živých organismů přitom
není dnes již vnímána
jako jakýsi samozřejmý a
mechanicky se odvíjející proces,
generující neustálý
„pokrok“
přiznává se role náhodě,
generátorem či akcelerátorem
evolučních změn
jsou často určité náhlé změny
a zvraty ve vývoji Země,
na ně musely živé organismy
reagovat, přizpůsobit se,
existovalo i několik
variantních řešení,
to, které se prosadilo nelze
pokládat za samozřejmé,
vyplynulo z určitých
konkrétních podmínek,
pokud by byly parametry
nastaveny trochu jinak,
mohlo se prosadit jiné řešení
sociologie zdůrazňovala
význam kultury –
v okamžiku, kdy vznikl
člověk, přestala prý u něho evoluční teorie platit
antropocentrický názor,
že člověk je něčím
výjimečným, co se z přírody vymyká –
až po nejrůznější kuriózní
teorie s mimozemšťany atp.
(von Däniken)
dnes se na studiu
antropogeneze podílí široký okruh oborů
z oblasti přírodních i
společenských věd
fyzická antropologie
(bioantropologie), resp. paleoantropologie
přímé prameny –
pozůstatky hominidů ve
stratifikovaných geologických,
resp. archeologických
vrstvách
doprovodné nálezy
paleobotanické a paleozoologické –
rekonstrukce původního
prostředí
posléze i artefakty –
tj. přímo upravené a
používané předměty – nástroje
nepřímé prameny –
biologická morfologie
(anatomie, histologie,
embryologie, fyziologie, genetika, biochemie, biofyzika,
etologie – nauka o chování živých systémů)
prehistorie, etnografie a
etnologie, psychologie, sociologie
poznání procesu antropogeneze
je
a zůstane ovšem stále
otevřenou problematikou –
dáno už samotnou
fragmentárností a různorodostí pramenů,
které neumožňují jednou pro
vždy rekonstruovat celý proces –
vždy zůstává řada možností
výkladu,
hledá se ten
nejpravděpodobnější za daného stavu poznání –
řada paradigmat – tj.
myšlenkových schémat – konstrukcí,
která ovlivňují
v kontextu dobových znalostí,
ale i mentality, ideových
východisek
do značné míry samotný
poznávací proces
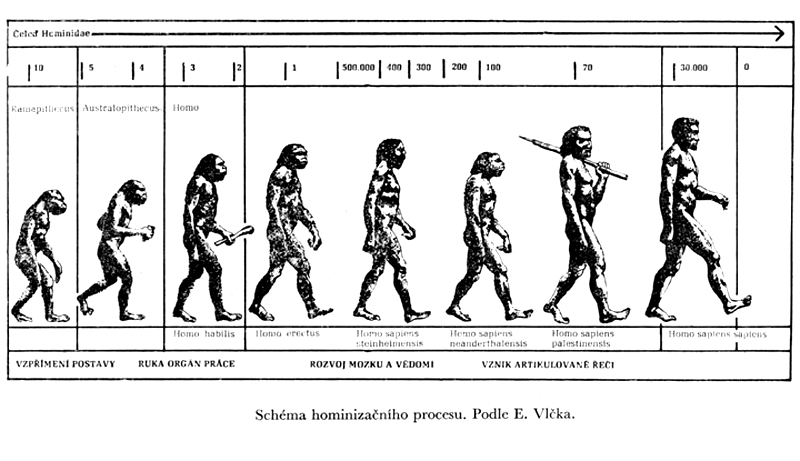
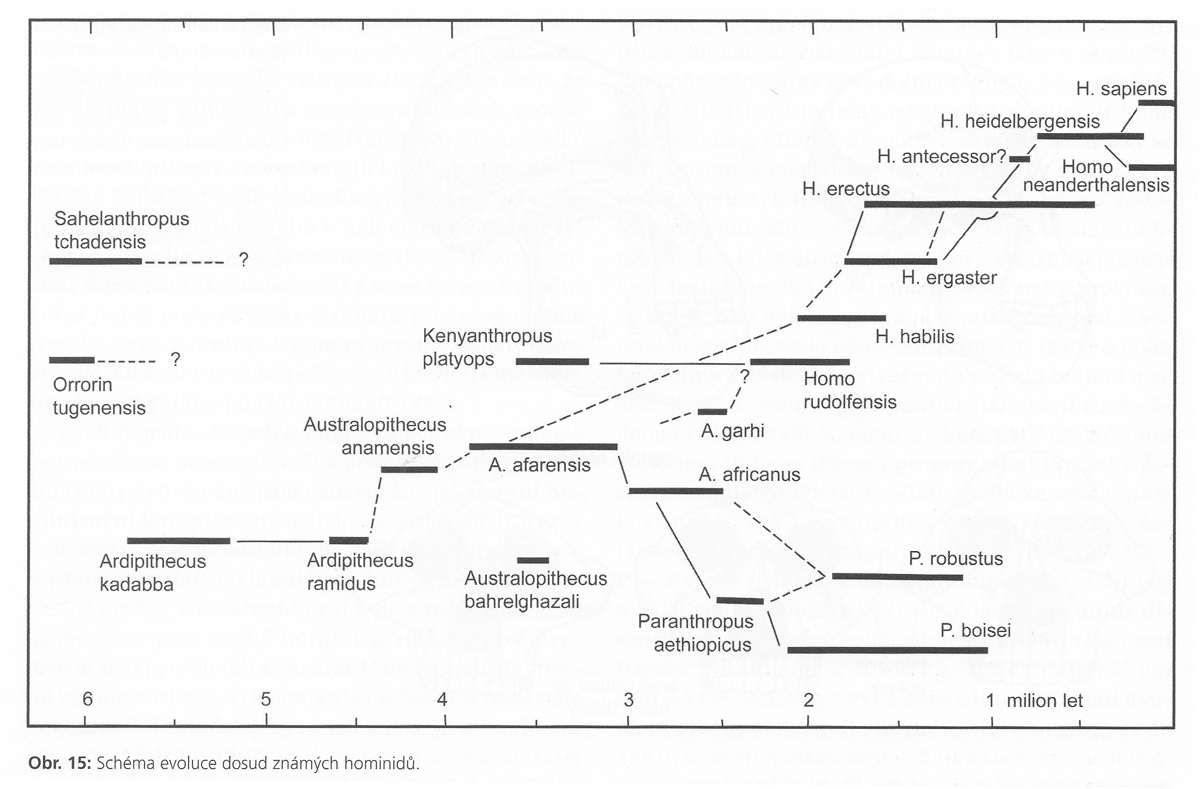
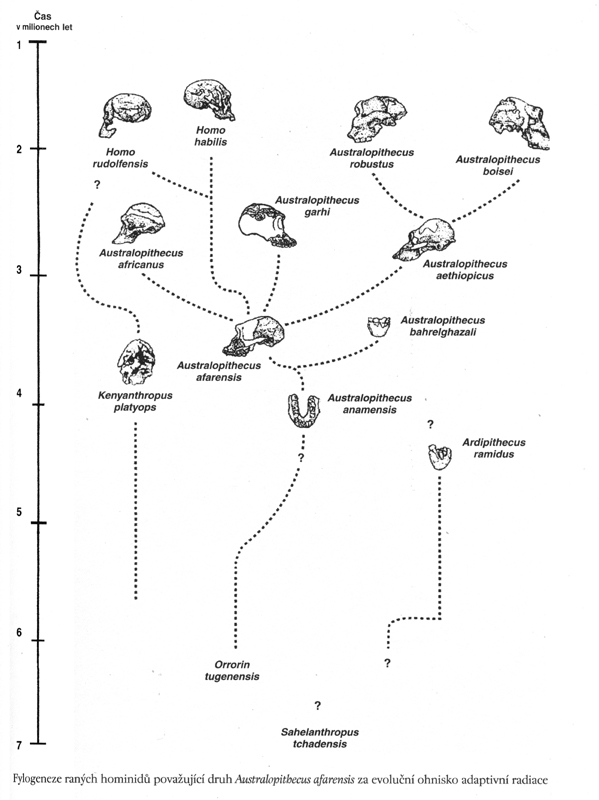
v rámci evoluční teorie bylo
např. dlouho rozšířené
paradigma víceméně lineárního
etapového vývoje:
australopiték –
pitekantropus – neandertálec – současný člověk
(platilo ještě v 60.letech
20. století)
další paradigma
70.-80.let připouštělo komplikovanější schéma
se slepými vývojovými články –
vyvinuté formy australopitéků
a později znovu neandertálci,
zároveň tendence hledat
počátky hominizace primátů co nejhlouběji v minulosti –
12-15 milionů let! –
již tehdy předpokládáno
oddělení od lidoopů, a to najednou –
formy – nejprve
Proconsul africanus (ten posléze prohlášen za lidoopí formu),
když objeven Ramapithecus
punjabicus a Kenyapithecus wickeri
(12-8 milionů let –
pliocén)
dnešní pohled –
např. bioantropolog Robert
Foley, Humans before Humanity 1995 –
česky Lidé před člověkem,
Praha 1998
aktuální český přehled:
Zbyněk Šmahel, Evoluce rodu Homo I-VI,
Živa 2003, s.37-40, 84-88,
134-136, 181-184, 227-231, 275-280
Václav Soukup, Dějiny antropologie, Praha 2004, II.díl, s.125-268
Viktor Černý a kol., Člověk v pravěku,
in: Archeologie pravěkých
Čech 1. Pravěký svět a jeho poznání, Praha 2007,
s.51-88
(zde i vysvětlení
k nejnověji užívaným metodám zkoumání lidských populací
rozborem DNA)
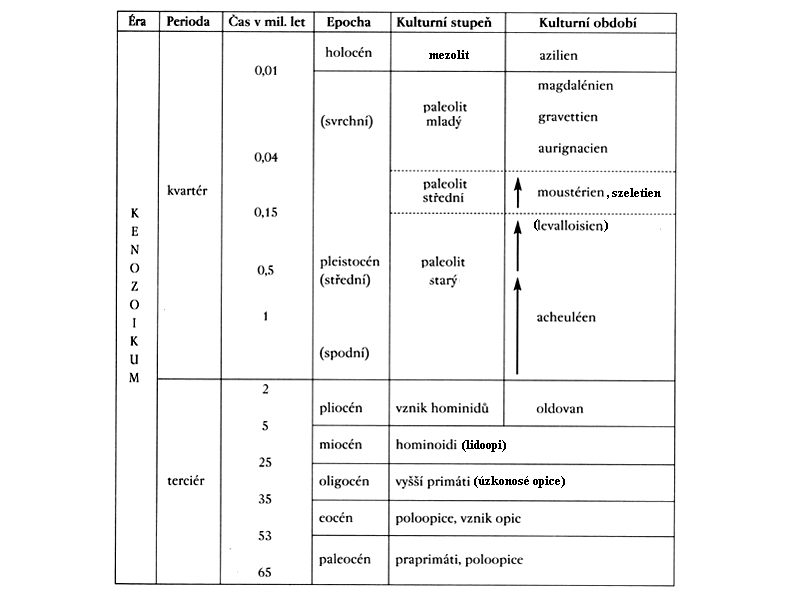
první savci – druhohory
- už asi 260 milionů let, dlouho ve stínu ještěrů
dinosauři vyhynuli asi před
60 miliony lety na konci druhohor –
katastrofa (srážka země
s velkým meteoritem?),
náhlá změna životního
prostředí, vyhynutí řady živých organismů,
naopak nástup těch druhů,
které se dokázaly se změnou nejlépe vyrovnat
třetihory – nejprve
jakési tápání –
nebylo vůbec tak jasné, zda
se po ještěrech prosadí jako dominantní
velké dravé formy nelétavých ptáků
nebo menší draví savci –
k vítězství savců
přispěl drift kontinentů –
mohli se nejprve nerušeně
vyvíjet v oddělené desce Asie,
kde velké dravé formy
nelétavých ptáků nebyly přítomny
a po propojení Asie
s ostatními kontinenty byli již draví savci
natolik vyspělí, že dokázali
ptáky zatlačit a likvidovat
posléze obrovský rozmach
všech forem savců –
obsadili prakticky všechny
ekologické niky na Zemi
řád Primates –
vývoj z drobných
stromových hlodavců –
od nejprimitivnějších poloopic –
ostrov Madagaskar - lemuři
orientovány ještě na noční aktivity
v dalším vývoji opice -
jisté rozvinutí pažního pletence, paží a vlastní ruky
=
uchování klavikul - klíčních kostí,
přestavba lokte umožňující pronaci a supinaci,
především pětiprsté končetiny s rozvinutou uchopovací
schopností
vlivem opozičního postavení palce,
nahrazení drápů nehty a rozvoj hmatových polštářků,
dále plochonohé dolní končetiny svědčící o alespoň
příležitostné bipedii,
frontálně orientované očnice umožňující prostorové
vidění,
redukce některých primitivních znaků obličejového
skeletu,
především změna zubního vzorce z I3-C1-Pm4-M3 na
I2-C1-Pm2-M3
(incisius = řezák, caninus = špičák, praemolarius =
třenový zub, molarius = stolička),
která přinesla zkracování obličejového skeletu,
regrese čichového laloku
a rozvoj koncového mozku společně se zvětšováním
mozkovny
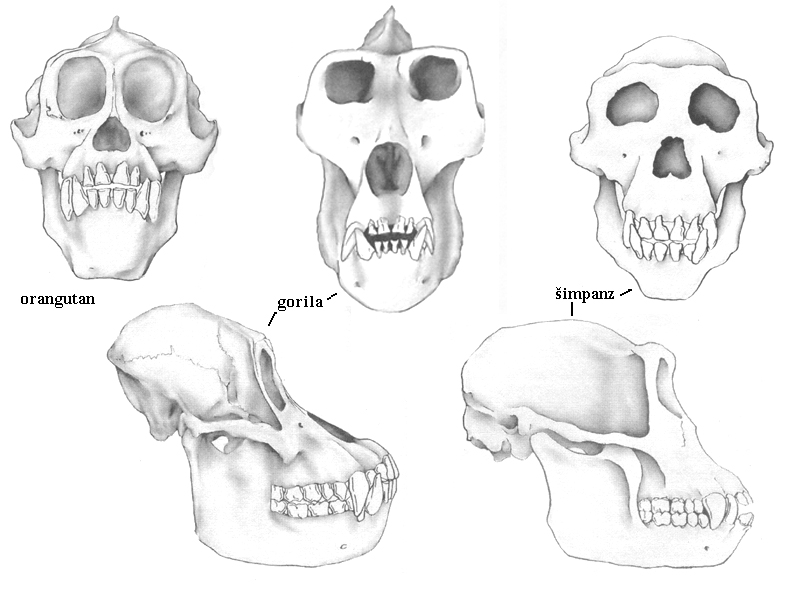
opice Starého a Nového světa
–
v Novém světě, tj.
v jižní Americe jen primitivnější ploskonosé opice –
dnes ohrožený druh
Amazonských pralesů
stereoskopické, ale barevně
ještě omezené vidění
(malý počet světločivých buněk
citlivých na světelné vlnění
v jednotlivých pásmech barevného spektra)
pokročilejší Starý svět
(úzkonosé opice) –
dokonalé stereoskopické
prostorové a barevné vidění –
důležité pro odhad
vzdáleností a pro výběr potravy (plody a ovoce)
dokonce i druhy,
které se dokázaly adaptovat
na prostředí mimo deštný tropický prales –
paviáni
– velmi vyspělá sociální organizace v samičích klanech
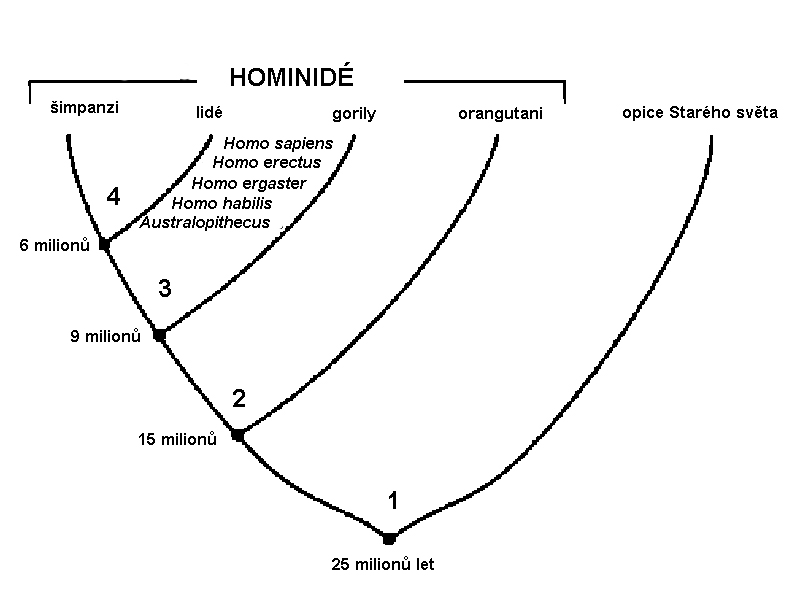
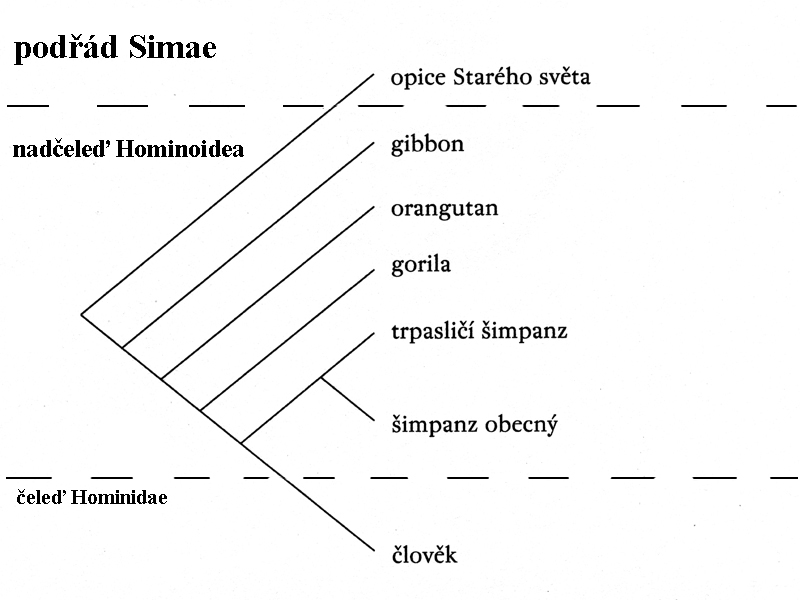
podřád Simae
(opice) –
nadčeleď Hominoidea (Anthropoides - nejvyšší primáti)
zahrnuje i dvě čeledi, které
již vykazují jisté lidské znaky.
Čeleď Pongidae s rody
dříve považovanými za lidské předky,
např. Aegyptopithecus 1, * 2
–
v oligocénu – asi
před 40 miliony lety –
zůstal ještě ocas, ale
proběhly závažné anatomické změny,
se znaky typickými později
pro lidoopy –
skelet je charakterizován
zpředu zploštělým hrudníkem
a rozvinutým pletencem
pažním, který umožňuje plný krouživý pohyb paže
před okolo 28 miliony lety
– tj. konec oligocénu –
počátek skutečného vzájemného
oddělení úzkonosých opic
a předků lidoopů (Hominoidea)
–
obě skupiny ve stejné životní
nice – tropický deštný prales,
opice spíše menší, pohybovaly
se po větvích po všech čtyřech končetinách
a využívaly k udržování
rovnováhy, ale i zavěšování ocas
lidoopi – zvětšení
rozměrů těla a váhy (přes 30 kg) –
pohyb v korunách stromů
spíše ručkováním – dlouhé přední končetiny
asi 25-12 milionů let –
miocén – Dryopithecus 1, * 2
či Sivapithecus 1, * 2 (Ramapithecus) – Indie
skelet vykazuje řadu pokročilých
„lidských“ znaků –
vlivem převažující bipedie, tedy chůze po dolních
končetinách,
dosahuje tento rod prahu prvního funkčního komplexu
procesu hominizace,
tj. dochází k přestavbě pánevního pletence
výskyt je datován asi 14 - 12 mil. let BP
(miocén/pliocén) –
nález ze Sivalické hory v Indii,
sdružování do tlup, užívání neupravených větví a
kamenů k obraně,
vegetariáni, příležitostná konzumace masa –
na procesuantropogeneze se však Ramapithekové
nepodíleli a vyhynuli
v miocénu pak započal
proces výraznější diferenciace mezi lidoopy 1, * 2 –
dáno hlavně geografickým
rozšířením
nejprve se oddělily formy
asijských lidoopů -
prapředci gibbonů
– asi 18 milionů let
pak orangutanů – asi
15-13 milionů let
skupina afrických lidoopů se
začala rozdělovat později:
až asi před 9-7 miliony lety
se oddělily gorily –
specializace na rostlinnou
potravu –
ke žvýkání listí a větviček
mohutné stoličky –
nezbytné neméně mohutné
žvýkací svaly –
úpony se sagitálním hřebenem
na lebce
výrazný pohlavní dimorfismus
–
vyplynul z uspořádání
sociálních vztahů (viz dále)
před 6 miliony lety se
oddělili šimpanzi –
poddruh šimpanz trpasličí (bonobo)
–
Z rovníková Afrika, lehčí
forma 35-40 kg, akutně ohrožen vyhynutím,
vlastní druh šimpanz učenlivý
(Pan troglodytes) – váha asi 55-60 kg,
pohlavní dimorfismus
relativně malý,
spíše všežravec, doplňkově
dokonce organizovaný lov na jiné savce –
opice guerézy či mláďata
paviánů
jednorázové
používání předmětů jakožto nástrojů
(ulomené větve –
k přitažení nedostupných předmětů,
kameny –
k rozlousknutí ořechů, větvičky na vybírání termitů a mravenců),
ale i zbraní – opět
klacky a kameny –
háží jimi po predátorech,
využívají i k zastrašování při vzájemných soubojích
vyvinuté formy komunikace
–
vcelku jednoduchá paleta
zvukových signálů, ale využívají také mimiky a gest
v zajetí experimenty
s učením znakové řeči –
schopni se naučit a používat
až cca 200-300 znaků,
dokonce (v omezenějším
výběru) znaky učit sami další generaci –
tedy již kulturní chování
dtto v přírodě –
provádění některých činností specifické pro jednotlivé tlupy –
(např. způsob jak
rozlousknout ořech)
učí mu opět i další generace
schopnost sebeidentifikace
(poznají se např. v zrcadle) atd.
šimpanzi tedy druhem
geneticky nejbližším člověku
komparace molekul DNA, tj.
genetického kódu –
98,4 % shod! z 33
milionů chromozomálních bází–
tj. menší rozdíl, než např.
mezi africkým slonem a slonem indickým!
šimpanz tak má
k současnému člověku geneticky dokonce blíže než ke gorile!
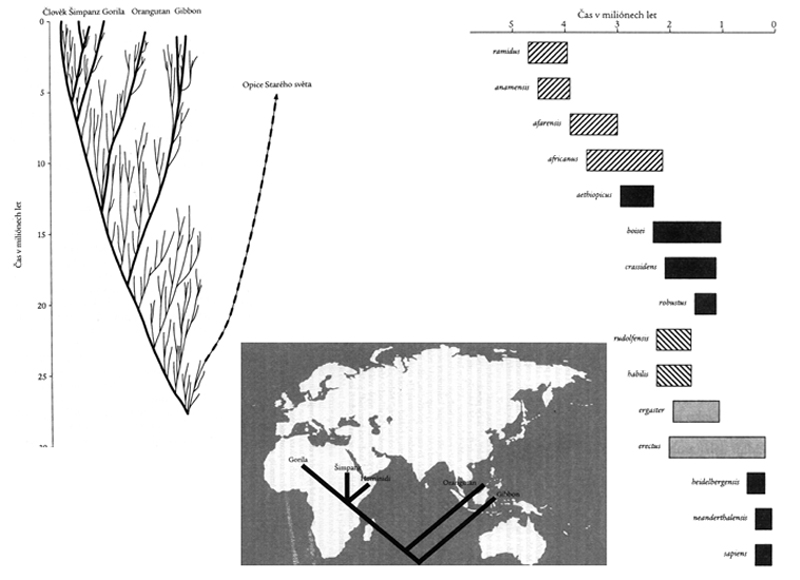
schéma
vzájemné příbuznosti druhů nadčeledi Hominoidea
i když samozřejmě čistě
statistické srovnání počtu rozdílů
v zápisu genetického
kódu je silným zjednodušením –
nevypovídá nic o rozdílech ve
významných sekvencích kódu,
které mají zřejmě podstatně
větší váhu
než shody v jiných méně
důležitých sekvencích –
z tohoto hlediska výše
uvedené srovnání vědci označují
za typicky zjednodušený
publicistický přístup
ke studiu šimpanzů
cf. Roger Fouts, Nejbližší příbuzní, Co jsem se dozvěděl od
šimpanzů o nás,
Praha 2000
pro všechny lidoopí formy
platí,
že vlastně od třetihor
nezměnily podstatněji způsob života,
zůstaly žít v původních
životních podmínkách deštných pralesů –
dokázaly se sice adaptovat i
na terén s rozvolněnější stromovou
vegetací –
kde lidoopi museli překonávat
větší vzdálenosti po zemi –
při chůzi po zemi ovšem
používají i dlouhých předních končetin – rukou,
o jejíchž zápěstí se při
chůzi opírají,
od původně spíše jen
rostlinné potravy přešli např. šimpanzi
k pestřejší skladbě
živin –
živočišné bílkoviny ale jen
v omezené míře –
adaptovala se také sociální
organizace lidoopů,
a to dost různorodým
způsobem:
úzkonosé opice – tzv.
samičí klany,
tj. tlupy tvoří společenství
příbuzných dospělých samic s mláďaty,
zatímco dospělí samci se
rozptylují a vyhledávají cizí tlupy samic,
k nimž se připojují jen
krátkodoběji
lidoopi:
gibboni –
žijí v monogamních
svazcích –
rodinu tvoří samec se samicí
a nedospělými potomky
orangutani –
žijí samotářsky, silní samci
mají několik samic,
které ovšem žijí individuálně
s mláďaty
a samci se k nim
střídavě přidružují (tzv. rozptýlený harém)
gorily –
silní samci si vytvářejí
harém samic,
mladí samci žijí mimo tlupu
nebo v podřízené pozici,
až uvolněním pozice vůdce
tlupy může nejsilnější ze synů
zaujmout místo po otci
harémové systémy sociální
organizace jsou zjevně příčinou
výrazného pohlavního
dimorfismu!
šimpanzi –
žijí v tzv. samčích
klanech,
tj. vytvářejí se menší a
dosti složitě strukturované tlupy o více samcích,
kteří mezi sebou soupeří o
dominantní roli v tlupě, tedy i o samice
systém, který nejvíce
stimuluje vyspělejší formy sociálního chování –
volba strategií, nezbytnost
uzavírat zájmové aliance s jinými jedinci atd.
závěr třetihor –
pliocén
východní Afrika –
po střetu litosférických
desek vyvrásnění vysokých pohoří v S-J směru -
tím zde dešťový stín, sušší
klima,
zmenšování rozlohy deštných
pralesů a přeměna v lesostep a savany
tomu se část populací lidoopů
musela přizpůsobit – a to velmi podstatně
bipedie – tj. orientace
na chůzi po dvou –
mění se stavba pánve a
dolních končetin
asi před 5 miliony lety
–
ve východní a severní části
Afriky –
specifické prostředí, členitý
terén,
řada populací patrně bez
větších vzájemných kontaktů,
relativně náhlé změny
životních podmínek –
to vyvolávalo relativně
rychlý průběh adaptace na prostředí –
postupně vzniká dosti široká
a pestrá řada lidoopích bipedických populací –
bohužel, zatím takřka žádné
nálezy z klíčového období 10-5 milionů let BP –
nejnověji z doby 7-6
milionů let BP nálezy,
které komplikují dosavadní
paradigma –
Sahelanthropus tchadensis toumai 1, * 2 nález z r.2001),
Orrorin tugenensis –
oba nálezy z Čadu
Ardipithecus ramidus kadabba
- Etiopie
bipední formy v lesnatém
prostředí a nejen na území Etiopie, ale také v Čadu!
tedy západně od předpokládané
pravlasti bipedických lidoopů ve východní Africe!
uspokojivé vysvětlení se
teprve hledá
změny prostředí jistě
významným faktorem,
ale reakce na ně dána také
možnostmi a předpoklady dotčených populací –
na vysychání a úbytek pralesů
reagovaly např. i úzkonosé opice
a vyvinuly se u nich stepní
formy s vyspělou sociální organizací –
pochopitelně kvadrupední -
paviáni
bipední lidoopi
vyvíjeli se po značně dlouhou
dobu 5-4 – 2 miliony let –
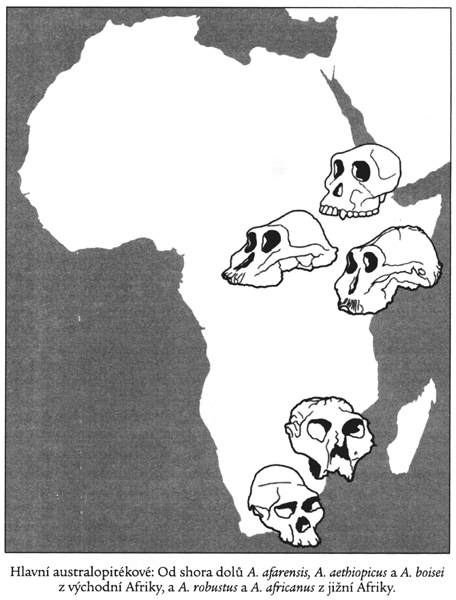
Australopitékové (jako rod čeledi Hominidae
= lidé)
rod Australopithecus však
zahrnuje řadu spíše lidoopích než lidských forem,
rané formy – 5-4
milionů let BP
Ardipithecus ramidus ramidus
(či správněji Australopithecus ramidus)
a Australopithecus
anamensis -
nový nález z Asa Issie
(leží 230 kilometrů
severovýchodně od etiopské metropole Addis Abeby) –
4,1 milionů let BP
dále Australopithecus
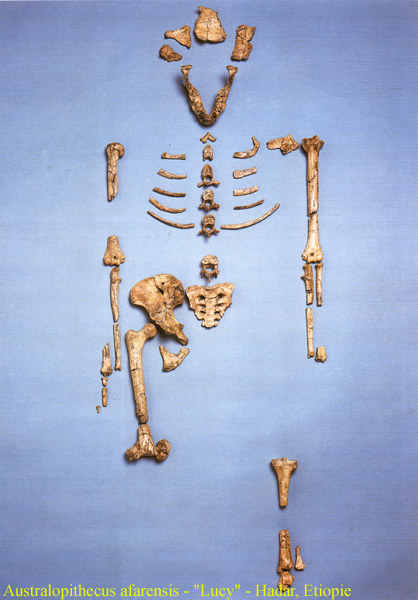
afarensis –4-3 miliony let BP
forma rekonstruována podle samičího skeletu z
Hadaru v Etiopii –
tzv. Lucy
– rekonstrukce lebky, stáří asi 3,1 milionu let BP
nález z r.1974 – jméno získala po tehdy
populární písni Beatles
Lucy in the
Sky with Diamonds
má zaoblený týl, nápadný torus supraorbitalis
(nadočnicový oblouk)
a bipední znaky ve stavbě pánve a femuru –
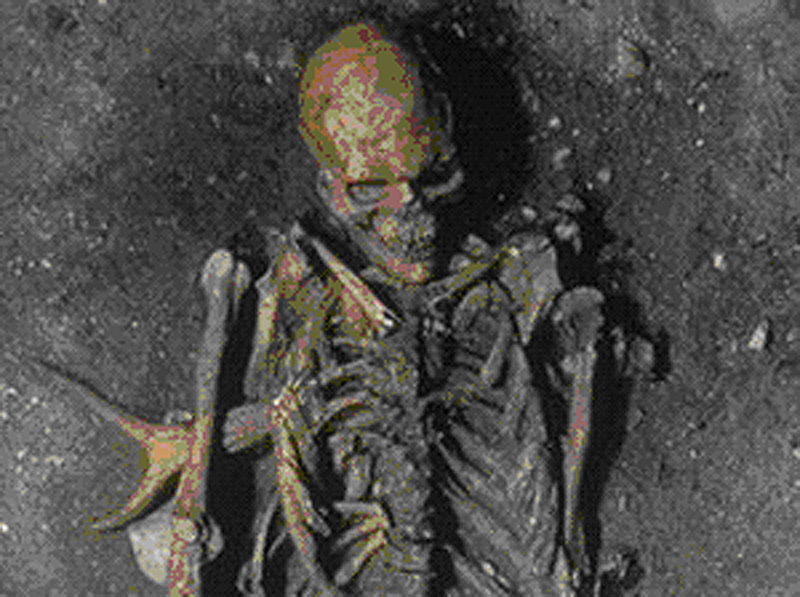
nový nález tříletého dítěte
(holčičky) u Dakaty v Etiopii,
o něco starší než Lucy – 3,3 milionů let BP
označena jménem Selam
zde vyobrazena lebka, ale dochovala se prakticky celá
kostra –
tedy mimořádně kvalitní nález,
umožňující zpřesnění dosavadních poznatků
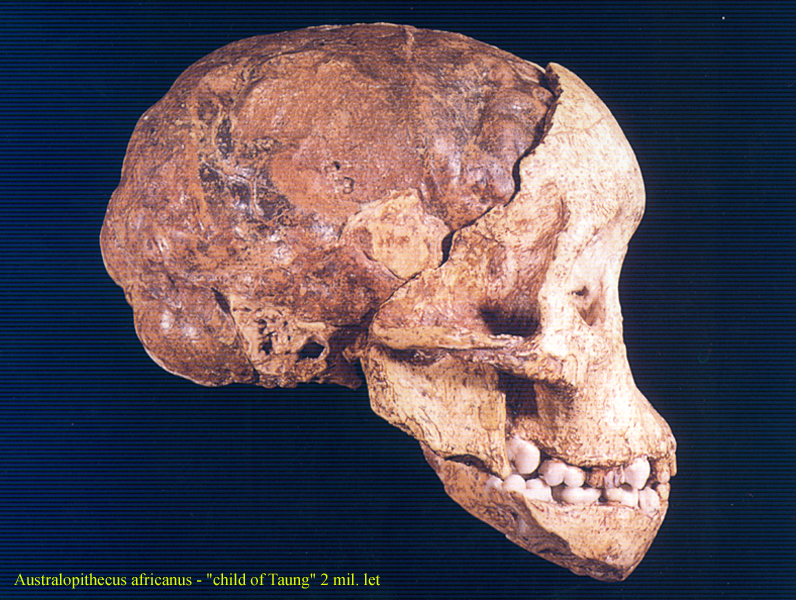
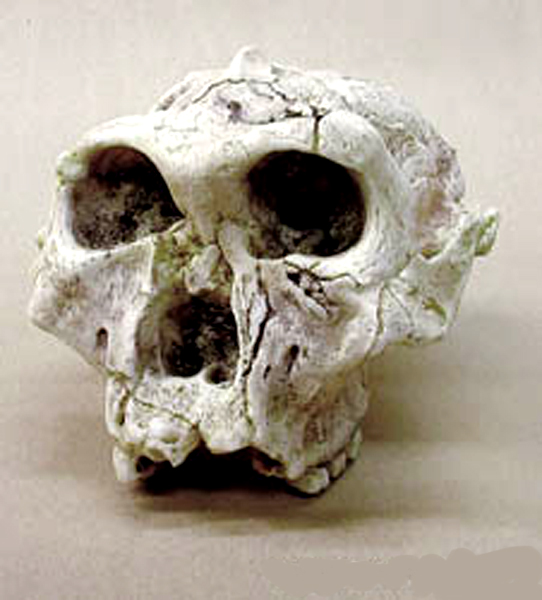
ještě pokročilejší forma Australopithecus africanus
–
3-2 miliony let BP – dítě
z Taungu – lebka 1, * 2
forma je ještě gracilnější, lépe uzpůsobená otevřenému
ekosystému
týlní otvor je dole, hlava je tedy držena zpříma, bez
vysunutí vpřed,
zuby nepřečnívají nijak výrazně, svědčí o častějším
příjmu masité stravy
používali kostěné, dřevěné i kamenné předměty,
ovšem bez záměrné úpravy či výroby -
Australopithecus africanus byl
proto některými badateli chápán
jako přímý předek zástupců rodu Homo
(v
současnosti ale převládá názor,
který za přímého předka
člověka pokládá pouze
starší formy Australopithecus
anamensis a afarensis
a od nich vedou vývojovou
linii již přímo k formám rodu Homo)
viz schéma
I
popsaní Australopitékové tedy spíše gracilnější
–
výška 130-145 cm, váha 35-45 kg,
nedokonalá chůze ve vzpřímené poloze,
ještě krátké stehenní kosti a málo vyvinutá klenba
chodidel –
(otiskly se do rozbahněného sopečného popela
a vzápětí překryty dalším popelem) -
dosaženo tak prvního fyzického komplexu hominizace,
který typický již jen pro Hominidae –
přestavba pánevního pletence a přechod k bipedii
výhodnost bipedie –
lepší výhled, horní končetiny osvobozeny od lokomoce
–
více využíváno jejich chápavosti k nejrůznější
manipulaci –
sběr potravy,
používání klacků a kamenů k obraně před predátory
vlivem vzpřímení dochází k posunu týlního otvoru na
spodinu lebeční,
dvojesovitému prohnutí páteře a vývoji meziobratlových
plotének
(kvůli mnohem většímu zatížení páteře a nutnosti
tlumení nárazů při chůzi),
vyklenutí a zesílení stehenních kostí,
přestavba kolene chráněného menisky, kotníků a rozvoj
nožní klenby
zároveň vzpřímení v dalším vývoji způsobilo
pokles hrtanu a prodloužení rezonační trubice
mozkovna nejvyspělejších
Australopitéků –
asi 400-550 kubíků (šimpanzi
mají obsah mozkovny asi 400-450 kubíků) –
v procesu antropogeneze
se pro porovnání velikosti mozku jednotlivých forem
používá tzv. indexu EQ
(encefalizační koeficient),
který ukazuje poměr mezi
velikostí mozku
a celkovou velikostí
(hmotností) organismu –
šimpanz dosahuje hodnot 2,5 EQ
–
většina australopitheků
rovněž
( jistou odchylkou u
progresivnějších forem směrem k 2,7-2-8 EQ)
čili intelektuální schopnosti
australopitéků nelze přeceňovat,
byly jen o něco málo vyšší
než u současných šimpanzů
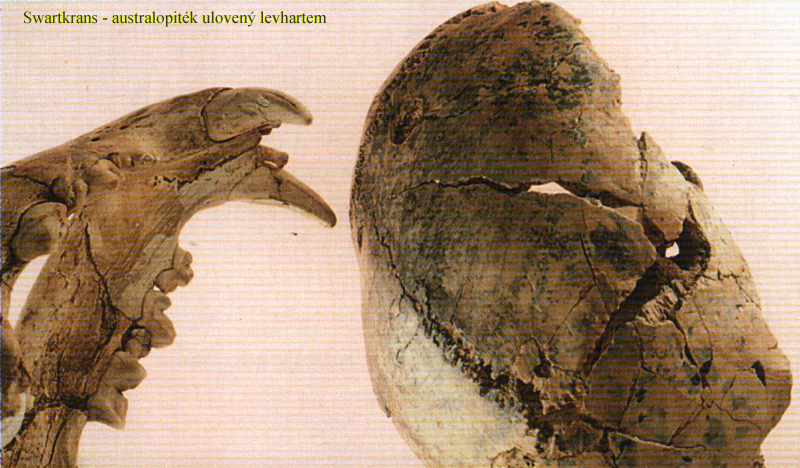
život v otevřenější
lesostepní krajině –
hrozba
predátorů – jedinci neobstáli,
ale koordinující tlupa
dokázala s užitím klacků a kamenů
levharta či šavlozubého tygra
zahnat
převaha rostlinné stravy -
sběr semen, hlíz atd.,
bílkoviny – hmyz,
vejce, lov drobných savců (myši a pod.) –
přísun hodnotných bílkovin
ty ale patrně získávány často
také konzumací mršin
život ve volné krajině
vyžadoval rychlý a vytrvalý pohyb,
spojený se značným
energetickým výdejem,
na který nebyly primáti
příliš vybaveni – přehřátí organismu! –
nutnost tepelné regulace
–
postupná ztráta srsti (vznik
„nahé opice“)
a zejména značné zvýšení
počtu potních žláz –
hominidé se výrazně více potí
než ostatní lidoopi
(člověk také označován jako
„potící se opice“)
další adaptace bipedických lidoopů
ve dvou liniích –
první z nich spočívala
v posílení tělesné konstrukce –
robustní formy, dosahující hmotnosti až 70 kg,
mohutný chrup, výrazný
sagitální hřeben na lebce (cf. gorily) –
umožňující úpon silných
žvýkacích svalů,
obličejová část tak výrazně
převládá nad mozkovou,
velikost mozku stagnuje,
původně se uvažovalo o lovu větších zvířat –
paviánů, kopytníků
dnes se zdůrazňuje naopak
skutečnost,
že zůstávají mohutné stoličky
pro rozmělňování rostlinné potravy –
i sagitální hřeben tak měl asi
podobnou funkci jako u goril
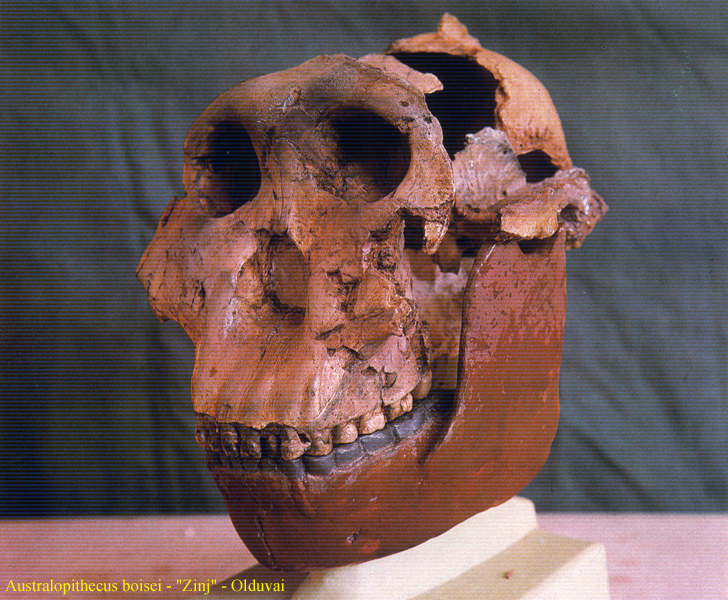
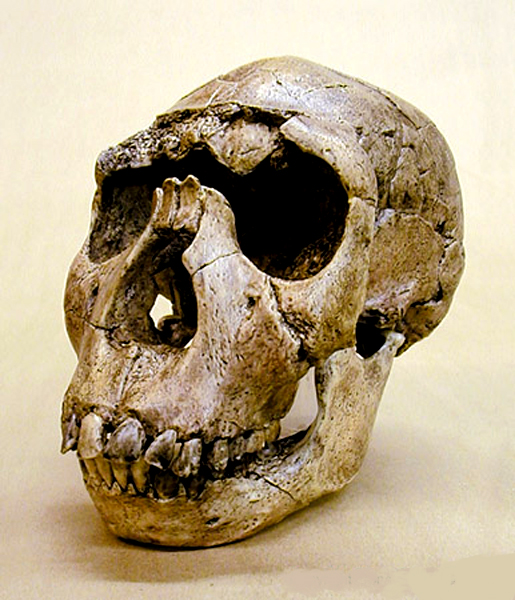
Australopithecus aethiopicus
3-2 miliony let BP
Australopithecus
boisei 1, *2
2,5-1 milion let BP
Australopithecus
crassidens 2-1 milion let BP
Australopithecus robustus 1,5-1 milion let BP
varianta vývoje hominidů
– schéma II
(porovnej s poněkud
odlišným pohledem na nejstarší genezi u schématu I)
vedle robustních
australopitheků ale vznikají formy,
které se daly jinou cestou
–
cestou výrazného posílení
sociability –
skutečné počátky rodu Homo -
vyvinuly se nejspíše
z formy Australopithecus afarensis
(uchovány vyčnívající špičáky
– typické pro všežravce, konzumující maso)
zlepšení kooperace mezi členy
tlupy - zvětšování mozku –
mozek je energeticky velmi
náročným orgánem, aby se mohl zvětšovat,
bylo třeba právě zvýšeného
přísunu hodnotných bílkovin –
první skutečně hominidní
formy: obsah mozkovny 640-800 kubíků! – EQ 2,9-3,1
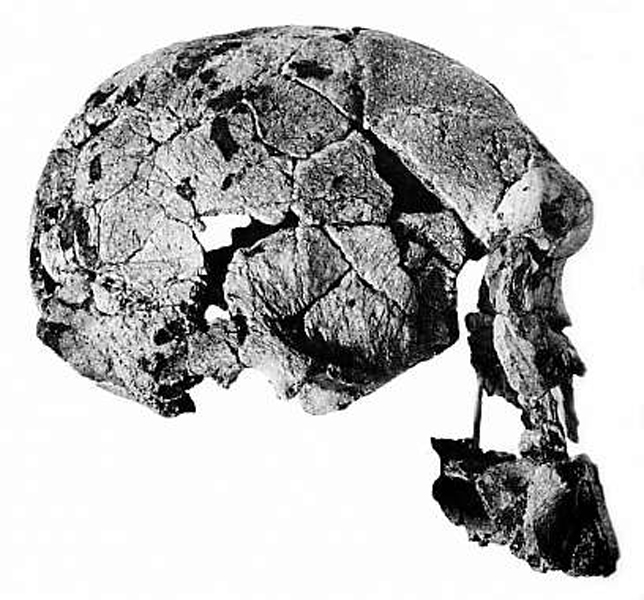
Homo rudolfensis – 2,5-1,7 milionu let BP (rod Homo)
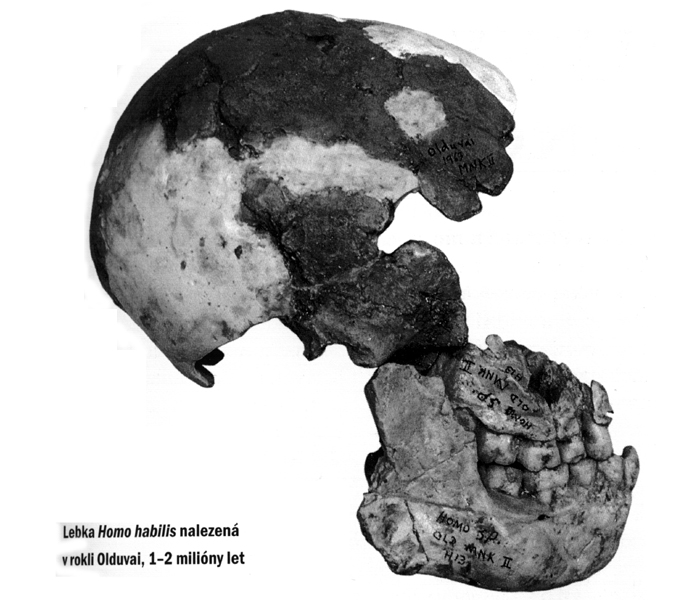
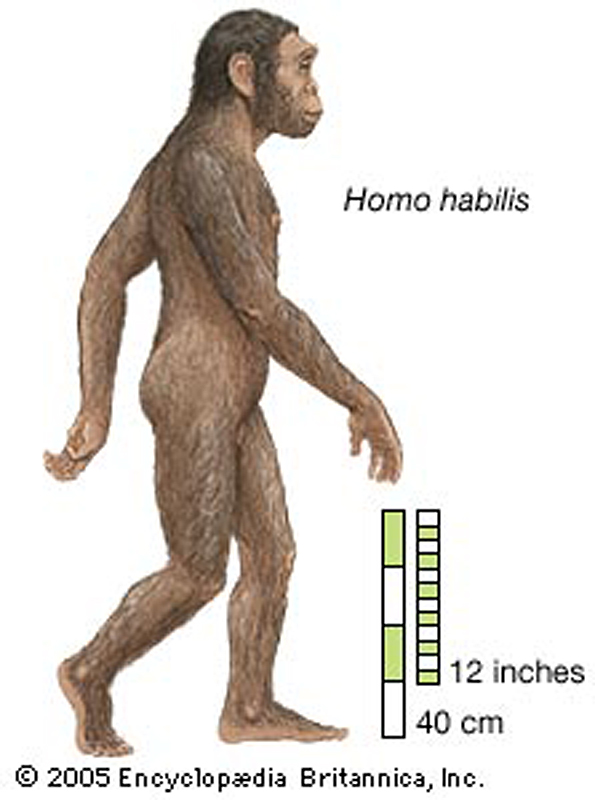
Homo habilis 1 , *
2 , * 3 – 2,3-1,7 milionu let
BP
žili tedy současně
s australopitéky, ale byli na vyšším stupni vývoje
první objev v Olduvaiské rokli v Tanzanii,
odkud pochází několik rekonstruovaných skeletů,
dále Turkana (Rudolfovo jezero) v Keni a lokalita
Gona v Etiopii
stále jistý sexuální dimorfismus –
muži výška 150 cm, ženy 120-125 cm, váha asi 40 kg,
nezbytnost kooperace v tlupách
však vylučuje harémové uspořádání sociálních vztahů
–
zůstává však zatím patrně zachována sexuální
promiskuita u žen,
spojená s výraznou sexuální rivalitou u mužů
–
prosazují se fyzicky nejzdatnější jedinci
nejvýraznější změny oproti australopithekům –
lebka –
celkově tenkostěnnější, vyšší, čelisti méně vystupují
obsah mozkovny tedy větší než 600 cm3,
chrup lidský se znaky všežravců,
výrazněji zaoblený týl lebky (vývoj týlního laloku),
naopak skelet se tolik od australopitheků neliší -
ruka - palec dosud krátký, prstní články široké a
zakřivené,
noha - prokazatelná úplná bipedie s vyvinutou nožní
klenbou,
ale dolní končetiny stále ještě krátké
tj. celkově tělesnými
proporcemi ještě stále blíže k lidoopům než k člověkovi
rekonstrukce
vzhledu, poměr velikosti lebky a mozku
doloženy jednoduché kamenné
nástroje – tzv. choppery (olduvajská
kultura),
tj. oblázky, u nichž úderem
bylo dosaženo
zatím spíše nahodilého
odštípnutí kusu kamene –
zbylý fragment oblázku tak
měl nějakou ostrou hranu či nepravidelnou špici –
to umožnilo daleko lépe
odřezávat a krájet maso, rozbíjet duté kosti (morek!) –
stále ale ještě nelze
uvažovat o náročnějším lovu větších savců –
ale naopak o rozvinutém
mršinožroutství –
důkazy – stopy na kostech
zvířat –
od zvířecích zubů a pak
překryty stopami řezání kamennou industrií!
jinak nepochybně jako
nástroje, resp. zbraně pro obranu -
dřevo – přiostřené
klacky či mlaty
či kosti
doloženo využívání ohně, i
když není jasné, zda jej už uměli rozdělat
sociální organizace –
přežití mohla garantovat jen
tlupa jako celek –
i ti nejzdatnější jedinci si
uvědomovali sounáležitost s ostatními členy tlupy
a dělili se s nimi o
potravu –
samozřejmostí v případě
těhotných a o malé děti pečujících žen,
jakož i nedospělých jedinců
vůbec – v nich perspektiva tlupy –
proto redistribuce potravy
– důležité pro další upevňování sociálních vazeb
rozšíření stále jen na
východní a severní Afriku!
současné paradigma –
formy Homo rudolfensis a Homo
habilis pokládá za přímé evoluční články
v procesu antropogeneze
jen s výhradou
(přímý podíl snad jen rané
formy Homo habilis) –
pak naopak chápány jako
dožívající vedlejší a slepé formy –
kolem 1,4 milionu let ještě
stále existovaly
vedle mnohem vyspělejší formy
Homo ergaster!
za novou progresivní formo
pokládán
okolo 2-1,9 milionů let BP a
až do 1 milionu let BP -
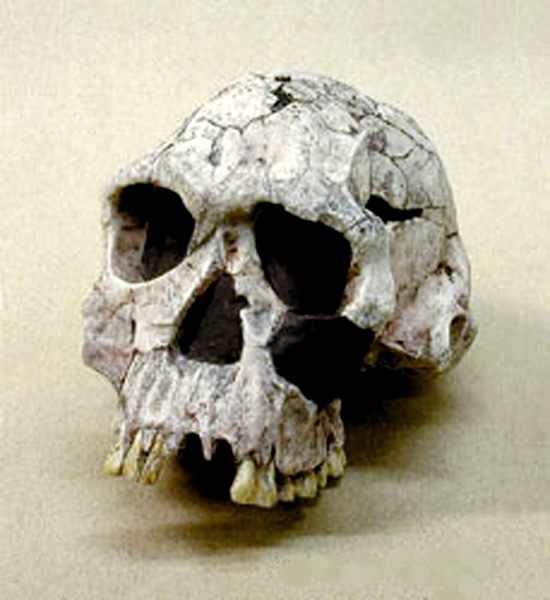
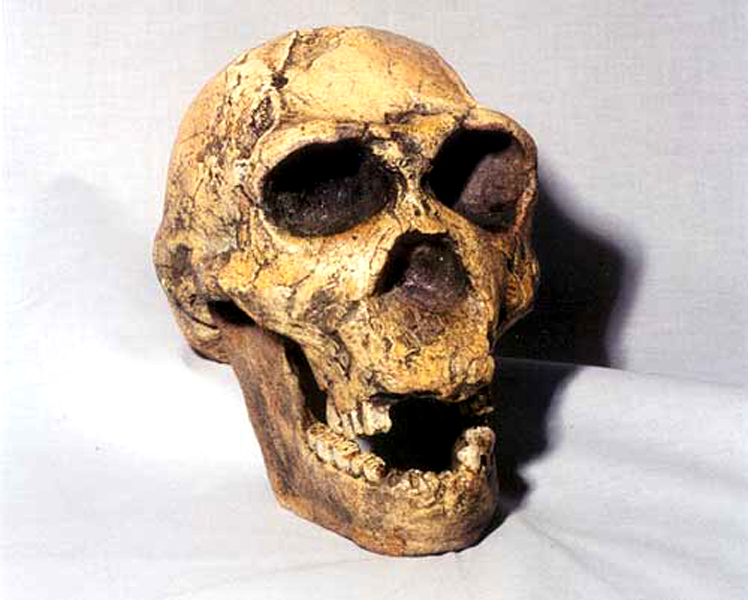
Homo ergaster
– Homo erectus
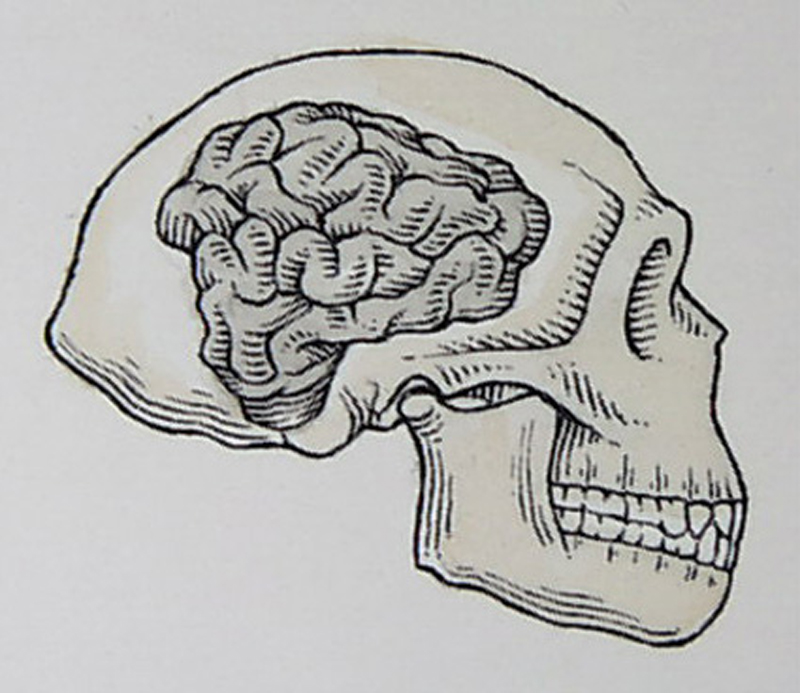
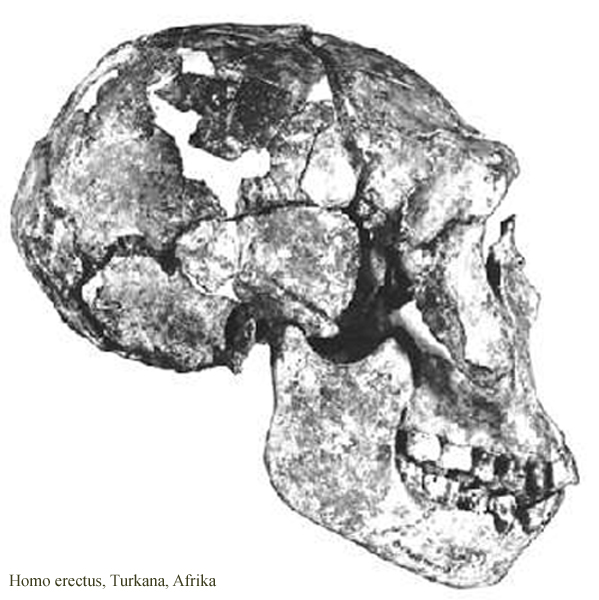
nálezy opět Turkana
v Keni – lebka 1, *2 –
lze demonstrovat typické znaky
–
silnostěnnější, mohutný
nadočnicový val, typické zalomení týlu
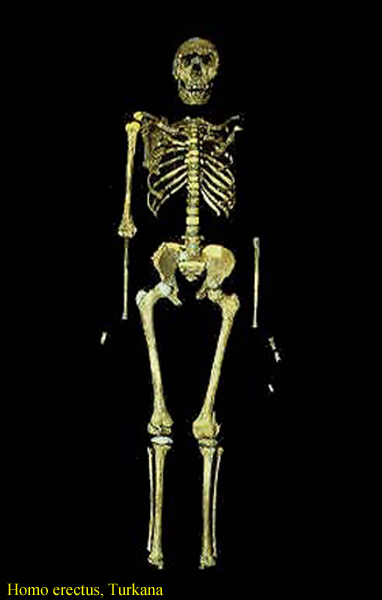
jiný skelet z Nariokotome – mladý muž
– lebka z Nariokotome
a ženská lebka z Koobi Fora v Keni
(aktuálně přestali badatelé
pro tyto africké nálezy
používat označení Homo
erectus, které vyhradili asijským formám)
kapacita mozkovny 700-900 cm3
– EQ až 3,5
objevuje se asymetrie
uspořádání mozku s dominantní levou hemisférou –
vyvíjí se tedy preferovaná
pravorukost! –
ruka patrně schopná jemnější
manipulace
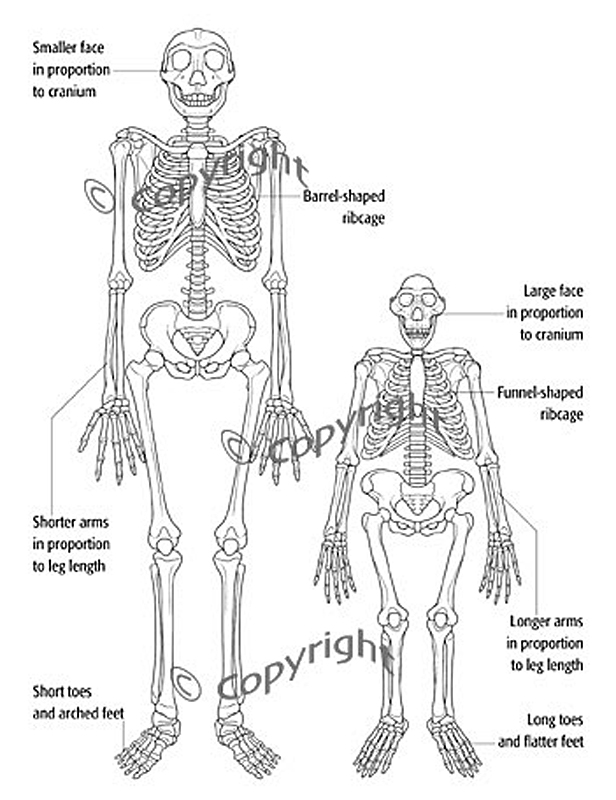
tělesná výška až 170 cm
zejména pak již tzv. lidská tělesná proporcionalita -
prodlužují se dolní končetiny
zmenšuje
se také sexuální dimorfismus
(pozvolný přechod
k párovým partnerským vztahům?)
vysoká postava –
rychlejší pohyb na větší vzdálenosti
větší tělesný povrch –
lepší termoregulace
v otevřeném a suchém tropickém prostředí
přechod od mršinožroutství
k aktivnímu lovu větších savců
zvýšený příjem živočišných
bílkovin z masa –
změny na chrupu – další
zmenšování stoliček,
naopak kvůli potřebnému stisku čelistí při ukusování
masité potravy –
vývoj nadočnicových valů
výše uvedená pravorukost a
zlepšení manipulačních schopností ruky –
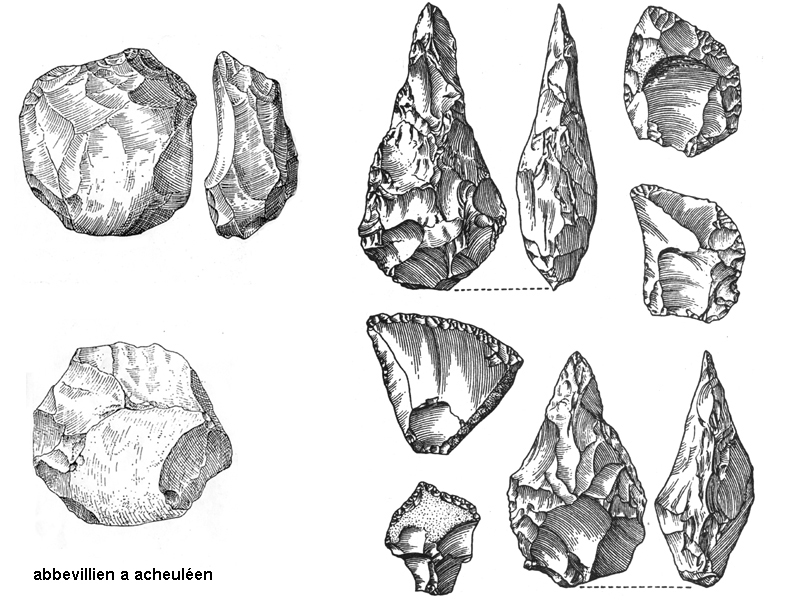
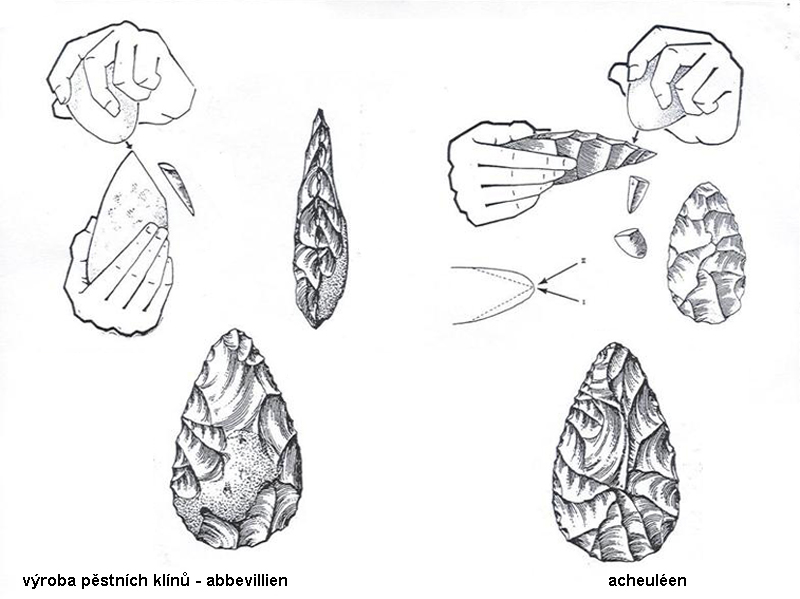
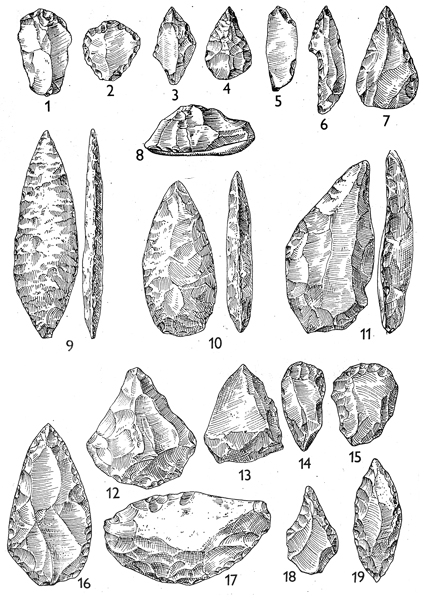
kamenná industrie – technologie výroby
nejprve tzv. abbevillien či cheuléen
(francouzská taxonomie
–
dnes se v češtině razí
používání přizpůsobené terminologie – čili cheulean) –
tj. industrie
z kamenných valounů (jader), již lépe opracovaných údery,
ale stále ještě zůstává část
původní kůry valounu (tedy navětralého povrchu)
ale kolem 1,7 milionu BP
z Afriky již nálezy industrie tzv. acheuleanu
např. Konso Gardula v Etopii
tj. opracován již celý povrch
kamenného jádra a vznikají tak nejstarší,
již pravidelnější formy tzv. pěstních klínů
srovnání
techniky výroby abbevillských a acheulských pěstních klínů –
ve druhém případě bylo třeba
silnějších a přitom přesněji vedených úderů –
náročné na koordinaci pohybu
rukou –
ale i myšlení - úderu musela
předcházet představa požadovaného výsledku!
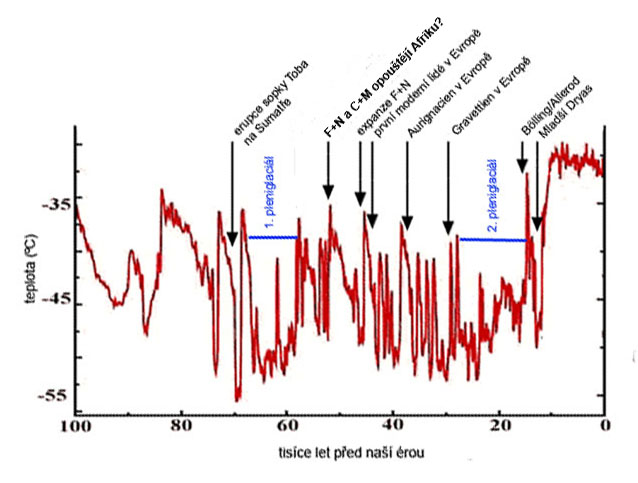
období okolo 2 milionů let BP
je také dobou expanze rodu Homo –
z Afriky do Azie
asi 1,9 milion let BP –
jeskyně Longupo v Číně, ale i Dmanisi v Gruzii –
patrně forma Homo ergaster,
která se v Asii rychle mění v typickou formu
tzv. Homo erectus –
charakteristické je menší
vyklenutí lebky, její prodloužení a zalomení týlu
jinak charakteristiky
zůstávají podobné formě Homo ergaster
expanze vyvolána opět
významnými proměnami životního prostředí –
okolo 1,8 milionu let BP
– počátek čtvrtohor –
celkové globální ochlazování,
neprobíhalo ovšem plynule
nýbrž pro celé starší čtvrtohory - pleistocén –
charakteristické je výrazné
globální kolísání klimatu –
doby ledové a meziledové
(glaciály a interglaciály),
v rovníkové oblasti
pluviály a interpluviály (vlhčí a sušší období),
označení glaciálů má
regionální platnost –
ve střední Evropě odvozeno od
rozšíření vnitrokontinentálního alpského ledovce
(Ginz, Mindel, Riss, Würm),
integlaciály značeny jako
Ginz/Mindel atd.,
řádově dlouhá období
glaciálů, v nichž klima opět dlouhodobě kolísalo,
lze členit jemněji na
chladnější stadiály a teplejší interstadiály,
v rámci posledního
glaciálu Würm rozpoznatelná i kratší klimatická kolísání –
mikroglaciály
příčiny klimatických změn
v pleistocénu –
patrně kombinace řady faktorů
(kolísání intenzity
slunečního záření,
změny polohy zemské osy vůči
ekliptice –
což při nerovnoměrném
rozložení pevnin a oceánů
vyvolávalo rozdíly
v bilanci přijmu a vyzařování tepelné energie)
důsledky klimatických změn
– výrazné posuny biotopů –
např. ve střední Evropě
v době vyvrcholení glaciálů věčně zmrzlá půda,
tundra, připomínající dnešní
podmínky u hranic polárního kruhu (severní Sibiř),
v nejteplejších obdobích
integlaciálů naopak až subtropické podmínky
zejména Dálný východ –
asi od 1,7 milionu let BP
jihoasijská tropická větev
Homo erectus - s vyšší postavou –
starší fáze Homo
erectus modjokertensis – Jáva (lokality Trinil a Modjokerto),
výška 170 cm, mozkovna 800 cm3,
asi od 800 000 let BP -
mladší fáze Homo erectus
erectus – mozkovna 900 cm3,
severoasijská větev –
nižší postavy (okolo 160 cm)
starší fáze Homo erectus
lantianensis – mozkovna cca 780 cm3
1,5 milionu let –
800 000 let BP
trochu nižší a méně hmotný,
takže EQ zůstává okolo 3,5
mladší fáze Homo erectus pekinensis – asi 700 -
200 000 let BP
Čína – jeskyně
Čou-kchou-tien u Pekingu
velikost mozkovny až 1050 cm3!
EQ se blíží k hodnotě 4
ale na Dálném východě dokonce
nověji nálezy Homo erectus
z doby asi 50 000
let BP
Jáva - Ngandong
dostal se dokonce do kontaktu
s formou Homo sapiens –
v konkurenci s ní
neobstál a vymizel
naposledy na indonéském ostrově
Flores
doložena gracilní až trpasličí forma Homo erectus
(výška jen asi 100 cm), která
vyhynula teprve asi před 18 000 lety
vysvětlováno jako výsledek
izolace od jiných populací
a života v nepříznivých
podmínkách s trvalým nedostakem potravy
jak již název říká –
Homo erectus měl vysokou štíhlou postavu –
výrazně lidských proporcí
lebka je silnostěnná
(patrně se tato vlastnost ve
studených klimatických obdobích
ukázala být výhodnou)
kulturní adaptace forem Homo
ergaster a Homo erectus na prostředí –
přestože zejména Homo erectus
žil mnohdy
v drsnějších
klimatických podmínkách, předpokládá se,
že dochází k další
redukci ochlupení –
mizí souvislý vlasový pokryv
těla –
naproti tomu schopnost
rozdělávat a udržovat oheň
pro vyhřívání obydlí a snad i
přípravu stravy
(efektivnější využití
potravin) –
převaha masité potravy
–
nezbytná právě k rozvoji
svalové hmoty
vyhledávání úkrytů pro
přebývání – jeskyně, převisy a jejich zateplování
organizovaný lov na velké
savce –
vyšší míra sociability
– schopnost myšlení –
tj. přehrávání akce předem –
v duchu –
tím uvědomování si příčin a
následků,
odhad jednání těch druhých,
nepochybně větší míra
komunikace –
ale patrně ještě ne mluvený
jazyk –
patro bylo ještě ploché a
velké,
byl ale již schopen vydávat
článkované zvuky –
tzv. předartikulační období
řeči –
dnes se zdůrazňuje také
význam gestikulace a mimiky v kombinaci se zvuky –
propojením vznikal systém
jakési znakové řeči –
v podobě zjevně mnohem
dokonalejší,
než jak ji dnes dokládají
experimenty se šimpanzy –
člověk byl již schopen tímto
způsobem
poměrně rozsáhlé a propracované
komunikace –
zvuky vyjadřoval pojmy, gesty
a mimikou naznačoval jejich vztahy –
tedy zárodek gramatiky
a tím již rozvoj abstraktního
myšlení
asijské formy Homo erectus
ale slepou vývojovou větví –
na Dálném východě přežívaly
velmi dlouho,
ale nepodílely se nijak na
vývoji moderního člověka a posléze vyhynuly
z Asie snad Homo erectus kolem 1 milionu let BP
do Evropy
a ještě později se snad zpětně dostal
v ojedinělých případech i do Afriky –
Ndutu – Keňa - 350 000 let BP?
v Africe ale odlišný vývoj –
od Homo ergaster přes méně výrazné erektoidní formy
již k formě Homo sapiens –
shodně s evropskou taxonomií používáno
pro nejstarší formy Homo sapiens v Africe
označení
Homo sapiens heidelbergensis nebo archaický Homo
sapiens –
EQ kolem 4
období mezi 1 milionem let BP až 200 000 BP však
poznáno velmi nedostatečně –
v tom slabina dnešních hypotéz
pak již zřetelný přechod k formě Homo sapiens
sapiens
opět ve východní části Afriky – od Etiopie
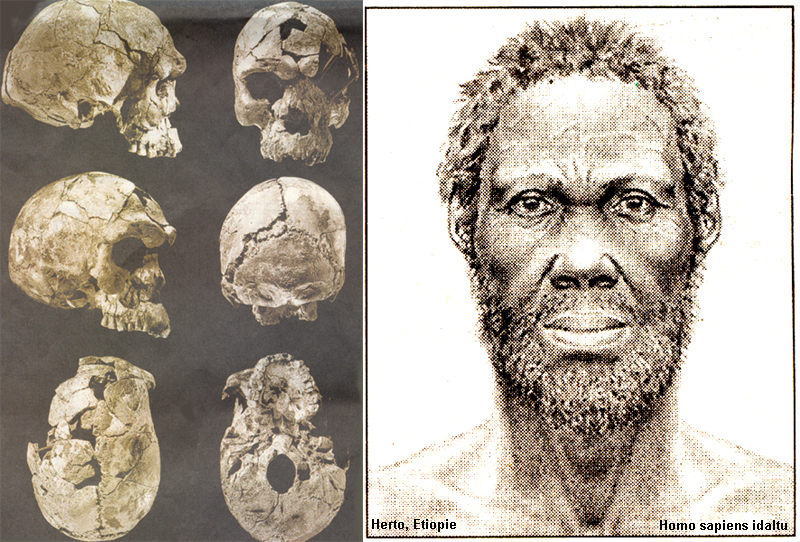
(Idaltu) po Jihoafrickou republiku –
Idaltu – Etiopie – Homo
sapiens idaltu – již okolo
300-280 000 let BP
chápán jako přímý článek přechodu
lebka s výrazně převládající mozkovou částí,
objem okolo 1500 cm3 ,
EQ tak roste na hodnoty okolo 5
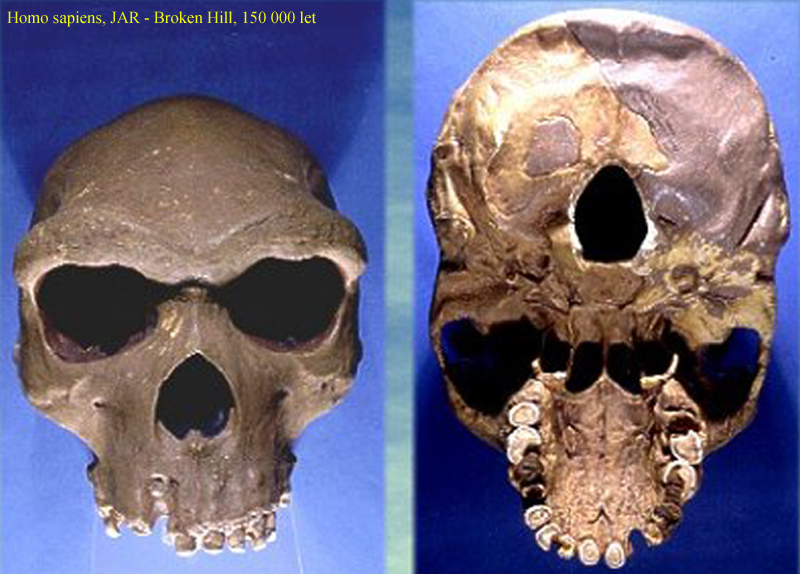
Zambie – Kabwe 1, * 2, *
3 (dříve Broken Hill)
ještě sice výrazné nadočnicové valy –
ale spodní čelist již s bradovým výčnělkem,
mění se postavení hrtanu – tím dostatečný
prostor pro hlasivky –
patrně přechod k artikulované řeči
mezi 200 000-140 000 BP vlastní Homo sapiens sapiens
mizí výrazné nadočnicové oblouky –
vpodstatě lebka současného člověka – až 1650 cm3
,
tedy více než dnes - EQ 5,3 ale jako u dnešního
člověka,
protože dnes sice kapacita mozku spíše mezi 1500-1550
cm3,
ale zase tendence ke gracilizaci postavy,
takže EQ zůstává v poměru k tělu na stejné
úrovni
Evropa –
po 1 milionu BP z Asie patrně Homo erectus
-
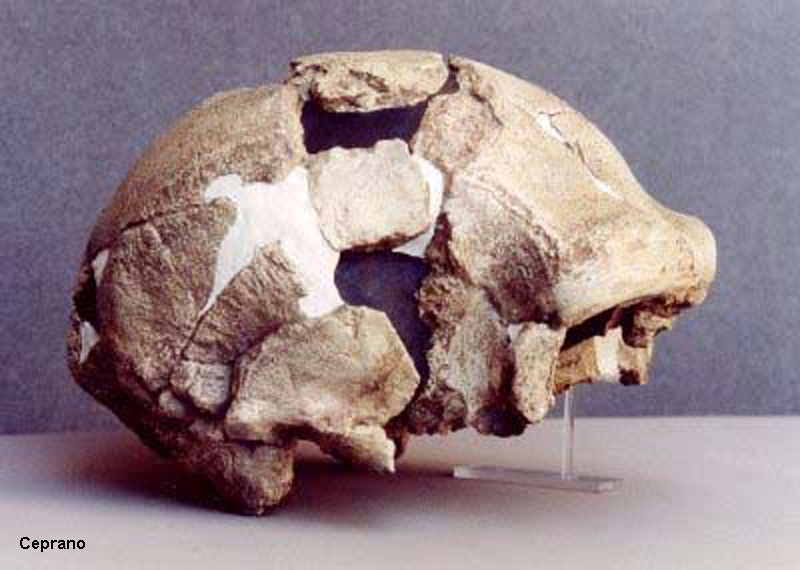
Ceprano – datace
900-800 000 BP
Mauer
a Bilzingsleben v SRN, Vértesszöllös v Maďarsku
velmi nejistě zatím část zubu z Přezletic
evropský Homo erectus byl poměrně robustní,
výška až kolem 180 cm, váha 80 kg,
mohutný trup a krátký krk,
mimořádně silná svalovina končetin a žvýkacích svalů,
dlouhá plochá mozkovna s výraznými supraorbitami a
obsahem až 1 100 cm3,
zmenšování obličejové části, zuby méně robustní,
čelisti ale značně vybočovaly a bradový výběžek nebyl
příliš vyvinutý
Homo erectus měl ještě širokou ruku s krátkými silnými
prsty,
jejichž pohyby byly poněkud toporné –
tím samozřejmě limitována úroveň vyráběných artefaktů
kulturně v Evropě – starší paleolit –
kultury cheleanská a acheulská –
definovány poprvé na základě nálezů z Francie
tedy primitivní kamenná jádrová, ale i úštěpová
industrie (odbíjením z jádra) –
clactonská či dokonalejší levalloiská technika –
hrubé pěstní klíny, či rovněž hrubé a masivní úštěpy –
Bilzingsleben a Vértesszöllös
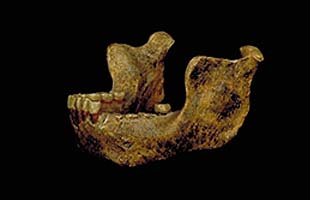
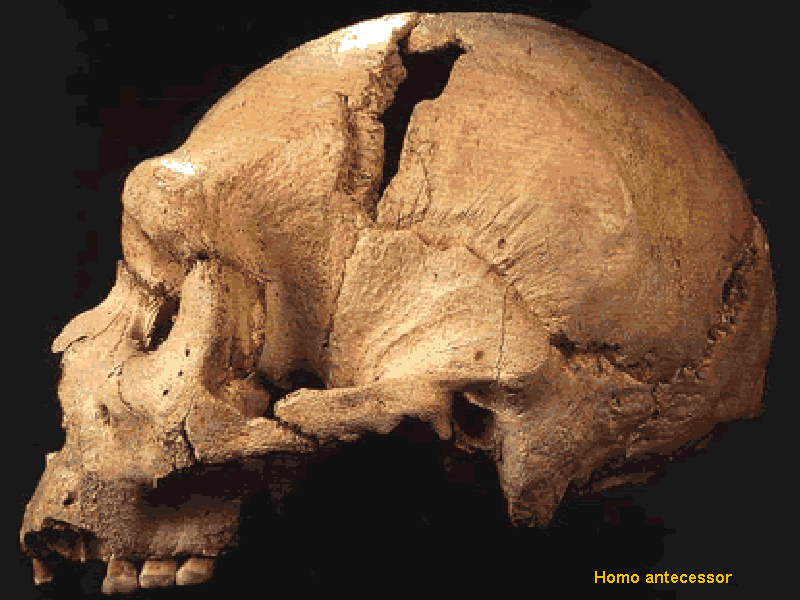
Grano Dolina – Španělsko datace cca 780 000 let BP –
forma nazvaná Homo antecessor
a rekonstrukce vzhledu –
doložen kanibalismus
některými badateli řazena k formám Homo sapiens
heidelbergensis,
které přišly do Evropy z Afriky,
jiní pokládají za samostatný vývojový článek –
na jedné straně souvislost s primitivnějšími
formami Homo sapiens heidelbergensis,
na druhé straně vyústění v pokročilejší,
byť slepý vývojový článek Homo sapiens
neandertalensis,
ale dokonce i možný přechod k formě Homo sapiens
sapiens
většina těchto nálezů datována až 500-200 000 let BP
–
tedy stále ještě starší paleolit –
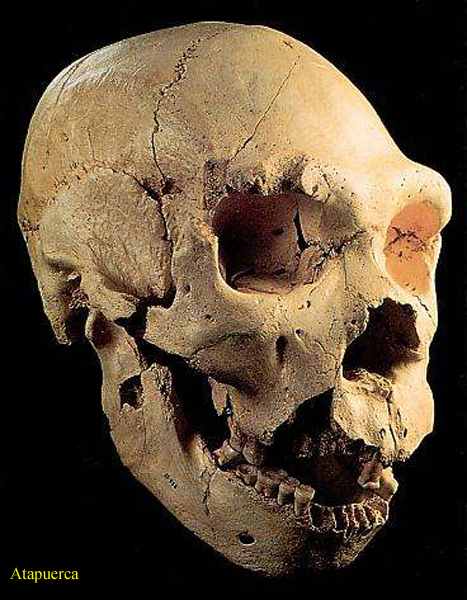
Atapuerca
ve Španělsku
(nálezy z Grano Dolina, Atapuercy a Sima de los
Hueos
pocházejí ze stejného území)
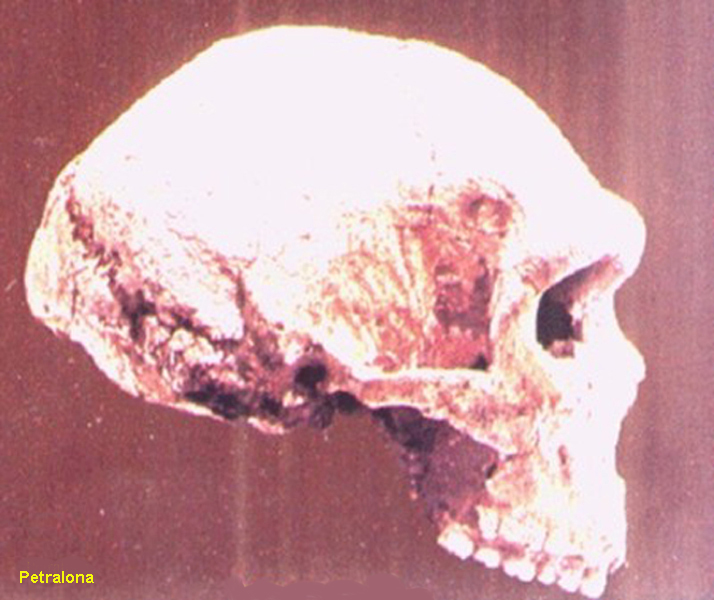
Petralona v Řecku
ta již některými znaky
naznačuje vývoj k formám
Homo sapiens neanderthalensis
–
nejstarší doklady asi
300-200 000 let BP –
Steinheim, Ehringsdorf,
Altamura – EQ 3,8 až 4,3
překvapivě variabilní populace ze Sima de los Hueos (Španělsko) –
32 jedinců z doby před
300 000 let BP
někteří nesou ještě starobylé
znaky Homo sapiens heidelbergensis,
jiní Homo sapiens
neandertalensis
a někteří dokonce vyhlížejí
velmi moderně
a blíží se tak formě Homo
sapiens sapiens –
celkově lidé velcí a robustní
– 180 cm, váha 80-90 kg –
s vyšší lebeční klenbou
a méně ustupujícím čelem
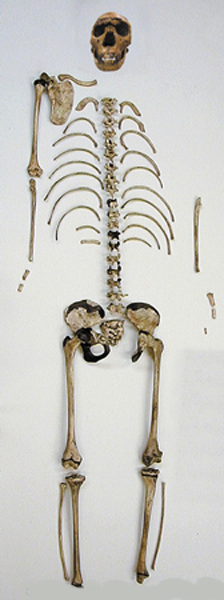
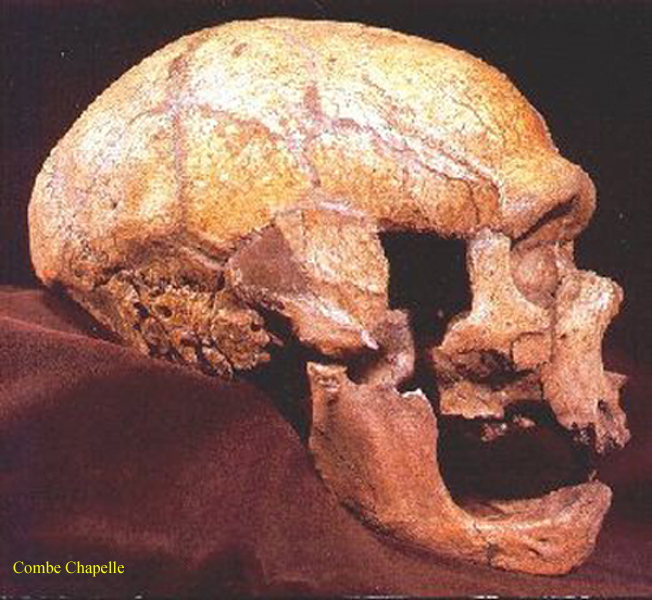
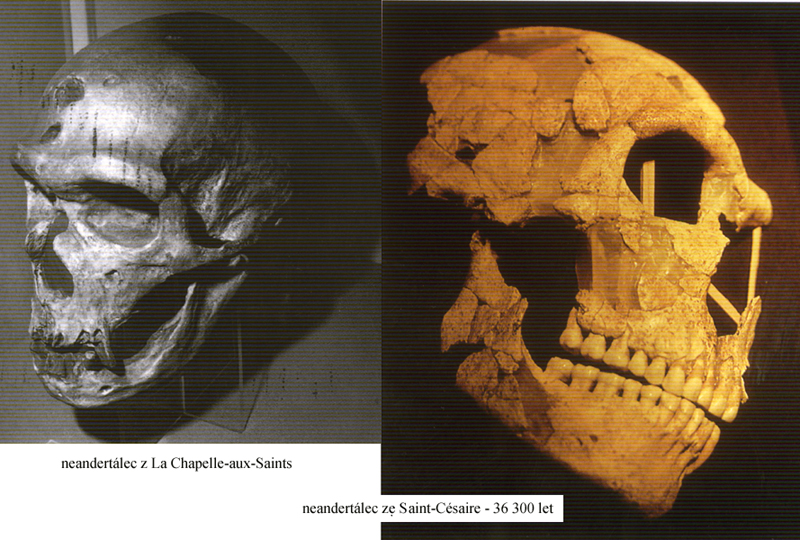
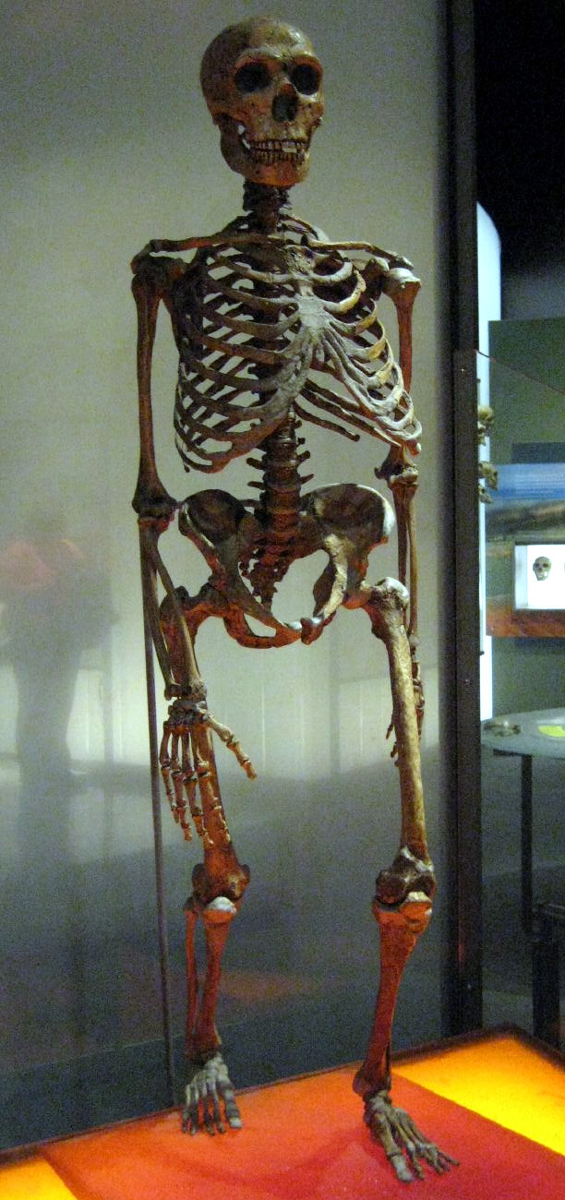
Homo
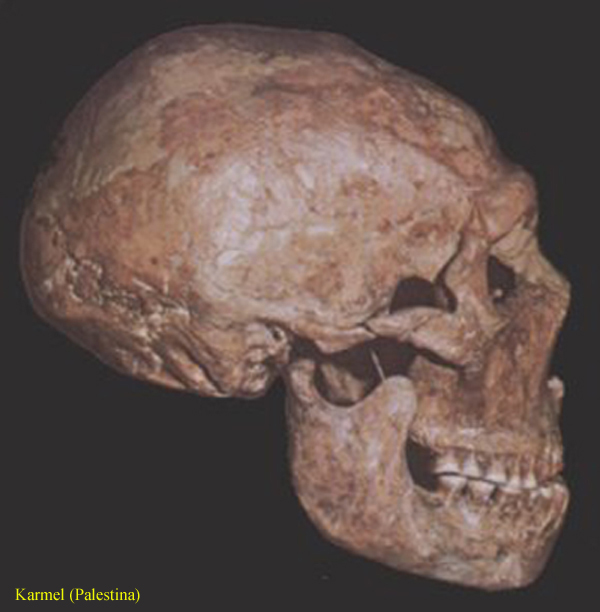
neanderthalensis 1, 2, – kostra
200 000 – 40 000 let
– tj. starší a střední paleolit – zejména glaciál Riss
v Evropě tehdy drsné
klima –
neandertálci formou dobře
adaptovanou na toto prostředí
celkově robustnější vzhled,
výrazné nadočnicové oblouky,
dolní čelist bez bradového výběžku
tento tzv. klasický
neandertálec ze středního paleolitu
(zejména z nálezů
v západní Evropě z doby před 100 000 – 40 000 let BP)
se stal zřejmě slepou
vývojovou větví
a byl v mladším
paleolitu vytlačen jinými, progresivnějšími populacemi –
robusností tělesných znaků se
odlišuje od ostatních forem,
končetiny jsou kratší, držení
těla nachýlené dopředu,
výška asi 165 cm muži, 153 cm
ženy
tedy nižší robustní postavy,
váha u mužů 80 kg
tím menší povrch těla –
menší tepelné ztráty
velká hlava, obsah mozkovny
dokonce někdy větší než dnes –
1 300 - 1 600 cm3
,
vzhledem k robustnosti
postavy ovšem EQ okolo 4,7
(tedy nižší než u současného
člověka)
lebka masivní,
s vystupujícími čelistmi,
mozkovna nízká, dlouhá,
s ustupujícím čelem a
prohnutým nadočnicovým valem,
na dolní čelisti dosud
chybějící či jen málo vyvinutý bradový výběžek
rozumová schopnost je
samozřejmě závislá také na kvalitě spojů,
nikoli jen na velikosti
mozku
pokročilé znaky na patře a
čelisti
i progresivní vývoj ruky nicméně
poukazují na vysoké postavení
v pomyslném vývojovém systému
jistá je schopnost nejen
udržovat, ale i rozdělávat oheň,
transportovat vodu a surovinu
v kůžích,
zřejmá je jistá ekonomie
surovin
a jistá míra prostorové
představivosti –
užívání levalloiské metody,
rovněž specializovaný lov,
sociální cítění (živení
zraněných a postižených)
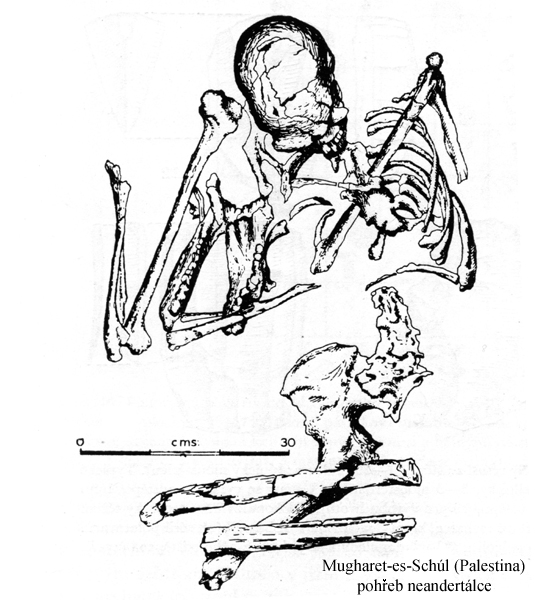
i vztah k mrtvým - pohřbívání jedinců –
první hroby, milodary nebo
doklady jistých představ –
body na kamenných deskách
kryjících hroby v La Ferraise,
květiny v Shanidar v Iráku
(zjištěno analýzou pylů)
nebo kozorožčí rohy v Tešik
Taš v Uzbekistanu,
nenutriční antropofagie
v jeskyních –
Krapina v Chorvatsku, Hortus
ve Francii, Guattari v Itálii
kanibalismus byl tedy
motivován magickými představami apod.
výlitek mozkovny klasického
neandertálce z Gánovců na Slovensku –
mimořádně významné pro
poznání vzhledu mozku –
členění mozkové kůry záhyby a
laloky
značný rozvoj
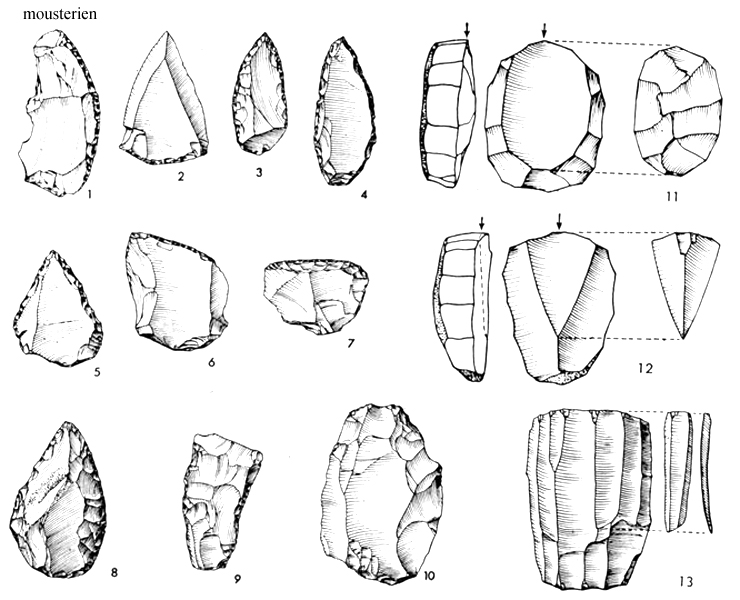
středopaleolitické industrie –
dokonalé pěstní klíny,
listovité hroty – kultury –
na 60 rozrůzněných typů
nástrojů
materiál – využívány
místní zdroje kamene –
tlupy patrně nemigrovaly na
větší vzdálenosti
lov velkých zvířat –
nosorožci, zubři, bizoni
v době meziledové
(Riss/Würm) sloni
ale i nebezpečných šelem
– medvědi
útok z bezprostřední
blízkosti – přímým kontaktem
lov vyžadoval nejen značnou
individuální sílu a mrštnost,
ale i taktiku, kooperaci,
vzájemnou komunikaci a pomoc
na kostrách časté doklady
úrazů
o zraněné a hendikepované
jedince tlupa pečovala –
doklady o zhojených těžkých
zraněních,
která by jedinec odkázaný sám
na sebe nemohl přežít
(nebyl schopen dlouhodobě
pohybu, zajišťovat si potravu atd.)
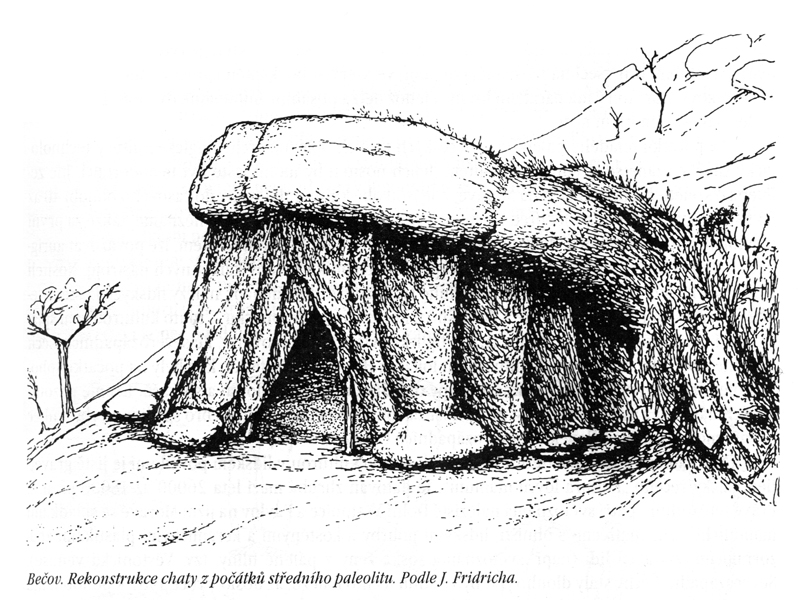
u nás – bezpečné
doklady pobytu až pro Homo erectus – Přezletice,
Bečov – Písečný vrch
– zde pak zejména střední paleolit –
získávání křemencové
suroviny,
patrně i chaty
–
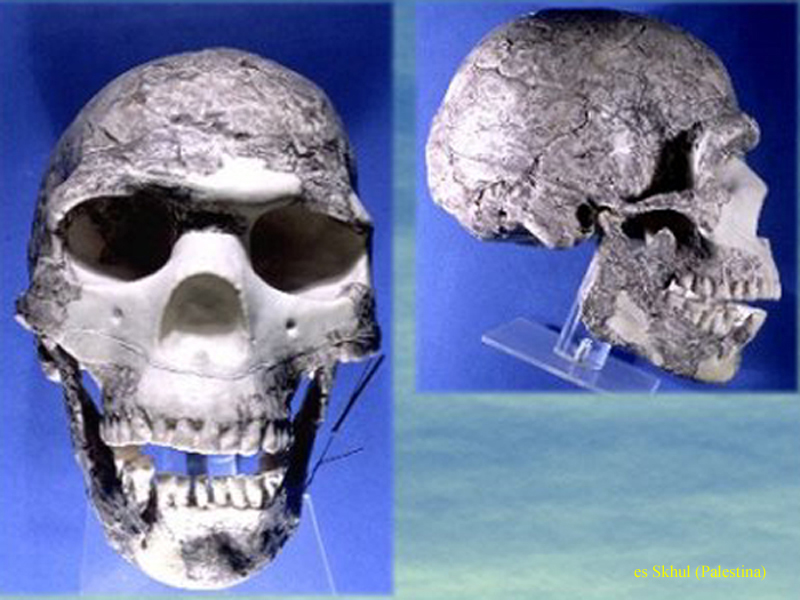
Blízký
východ 1, 2 – rovněž neandertálci, ale ne tak
vyhraněné formy
a zřejmě možný přechod
k populacím Homo sapiens sapiens
u Homo sapiens proběhly nejprve změny
tzv. třetího fyzického komplexu sapientace –
především na lebce - neurokranium je již větší než
splanchnokranium,
ústup supraorbit (nadočnicové oblouky),
rozvoj bradového výběžku, na nějž se váží úpony jazyka
a posléze i čtvrtého fyzického komplexu –
další rozvoj schopností manipulace vlastní ruky
a vznik schopnosti artikulované řeči –
oboje se váže na rozvoj levé mozkové hemisféry,
která ovládá jak zpravidla lépe k manipulaci
uzpůsobenou pravou ruku,
tak mluvidla
zmíněné prodloužení rezonanční trubice umožnilo větší
variabilitu tónů
a vystoupání patra
společně se zkrácením zubního oblouku, uvolněním rtů
a vystoupáním jazyka
z tzv. opičí jamky podpořilo –
také díky úponům na rozvíjející se bradě - artikulaci
zároveň se rozvíjí Broccovo centrum řeči v oblasti
spánkového laloku
a principy chápání a paměti, které jsou pro vznik
řečového kódu nutností
vyvinutí neandertálci měli ruku již obratnější, roste
délka palce,
hlasivky mají dosud ostré okraje
a bradový výběžek není příliš rozvinutý,
tvrdé patro je však již mírně vyklenuté,
předpokládá se drsný, hlubší hrdelní hlas s počátky
artikulace –
tzv. rudimentální článkovaná řeč
osud neandertálců –
dnes objasněn poměrně spolehlivě výzkumy DNA –
studium mitochondriální DNA – z mitochondrií,
tj. útvarů v buněčném obalu, které buňkám
zajišťují „výrobu“ energie
tato DNA přenáší se pouze prostřednictvím matky
a lze tak sledovat nahodilé genetické mutace
a vymezit, ve kterých populacích se vyskytují
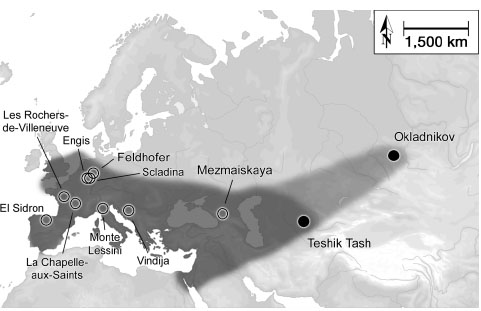
odhad populace nejvýše cca 70 000 neandertálců
na území od
Španělska po Sibiř
dohledáváním příbuznosti jednotlivých genetických
linií zpětně do minulosti
a odhadem časového průběhu změn bylo možné dospět
až k fiktivní pramatce celé současné lidské
populace -
tzv.
mitochondriální Eva – žila asi
před 200 000 lety v Africe,
Homo sapiens sapiens na celém světě vznikl výhradně z této
africké populace
rozložení populací Homo
sapiens sapiens ve světě
na základě analýzy mitochondriální DNA
genetické studie založené na mtDNA
(mitochondriálních) haploskupinách jsou méně
informativní
a mnohem „rozmazanější“,
než studie haploskupin mužského chromozomu Y,
protože ženské geny z řady důvodů „tekly“
mezi geneticky cizími populacemi
dtto studium genetické informace na mužském chromozomu
Y
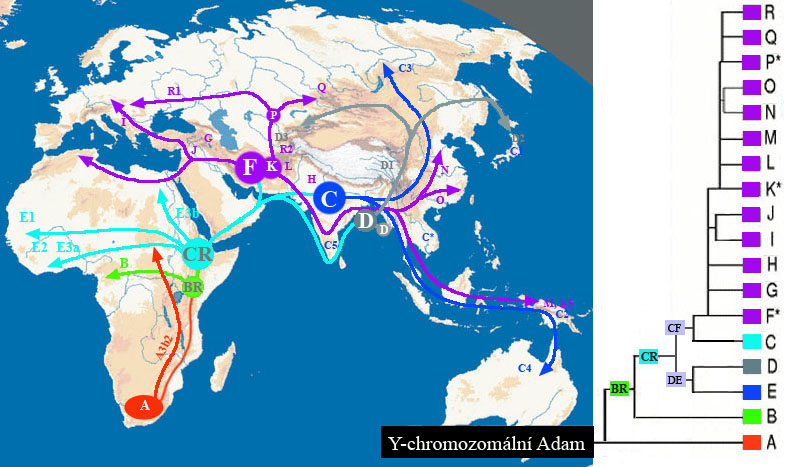
poloha M42 s archaickou variantou molekuly
adeninu –
zastoupena u lidí v omezeném počtu jen
v některých afrických populacích –
jako dědictví raných forem Homo sapiens
všechny ostatní populace pouze varianta
s molekulou thyminu –
tato mutace vznikla až v době formování
moderního člověka Homo sapiens sapiens
v jižní a východní Africe
a tato populace ji rozšířila po celém světě
a to opět mezi 200 000-140 000 lety – tzv. chromozomální Adam
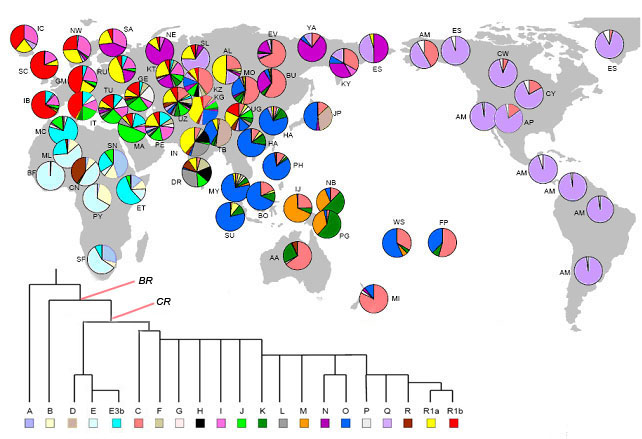
průběh rozšíření Homo sapiens
sapiens po světě
na základě analýzy mužského chromozomu Y
v malých tlupách doby kamenné docházelo snadno ke
genetickému driftu
(převládnutí linií určitého dominantního člena)
staré Y-haploskupiny se tudíž dnes vyskytují
v geograficky dobře vymezených oblastech,
které ukazují na místo jejich původu
protože muži jen zřídkakdy opouštěli vlastní klan,
Y-haploskupiny velmi pomáhají při zkoumání
historických migrací
rozdíly v DNA natolik velké, že lze předpokládat,
že k oddělení současného člověka od společných
předků
s Homo sapiens neandertalensis došlo už před
600 000 lety BP
(tedy v době Homo sapiens heidelbergensis)
neandertálci se tedy na vývoji současné lidské
populace prakticky nepodíleli –
nedošlo tedy k jejich asimilaci –
neexistují potomci vzešlí ze spojení jedinců obou
druhů,
kříženci totiž zůstávali neplodní
(podobně jako při křížení oslů a koní – mezkové
a muly se dále nemnoží)
přestože žili na Blízkém východě
a poté i v Evropě po nějakou dobu vedle sebe
neandertálci tedy v konkurenci
a leckdy patrně i přímé konfrontaci s Homo
sapiens sapiens
neobstáli a vyhynuli
poslední doklady existence – Gibraltar –
okolo 28 000 BP –
až moderní lidská forma Homo sapiens sapiens
je schopna jemné práce rukou –
umělecké předměty, technologicky náročné výrobky
–
a také plně artikulované řeči
tzv. nostratického či preboreálního jazykového
společenství
(k jeho štěpení tedy dochází až v průběhu
mezolitu?)
ukončení vývoje formy Homo sapiens sapiens
v Africe
asi před 100 000 lety, postupně se pak šíří,
asi před 40 000 lety
ovládla i Evropu a Asii,
odtud pak v závěru poslední doby ledové i do
Ameriky
kulturní vývoj v mladším
paleolitu – viz další téma
na závěr ještě jednou schéma antropogeneze
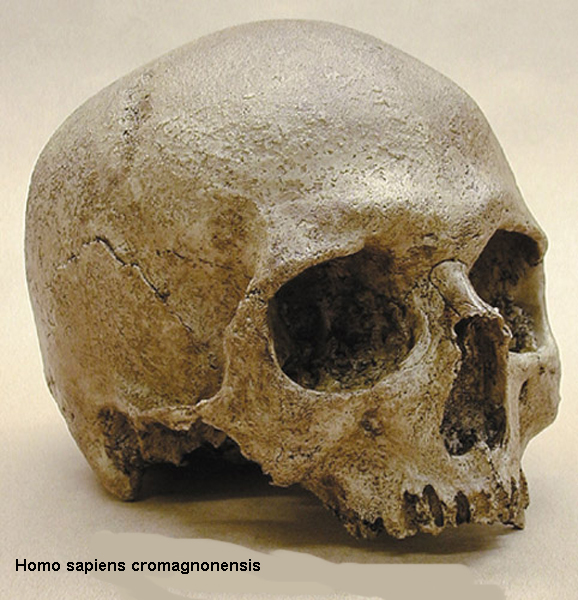
mladší paleolit (40-12 000
let př.n.l.) –
již jen Homo sapiens sapiens
- člověk cromagnonský, grimaldský atd.
relativně rychlé rozšíření po
celém světě, vznik dnešních ras
mozkovna průměrně 1400
kubíků, EQ 5,3
stavba těla jako současný
člověk, jen celkově ještě trochu robustnější,
hlavně ale rozvoj mluveného
jazyka, což značně urychlilo proces myšlení
Základní literatura:
Šmahel, Zbyněk, Evoluce rodu Homo I-VI, Živa 2003,
s.37-40, 84-88, 134-136,
181-184, 227-231, 275-280
Soukup, Václav, Dějiny antropologie, Praha 2004, I.-II.díl, s.15-268
Foley, Robert, Lidé před člověkem, Praha 1998
Fouts, Roger, Nejbližší příbuzní, Co jsem se
dozvěděl od šimpanzů o nás,
Praha 2000
Diamond, Jared, Třetí šimpanz, Praha - Litomyšl 2004
Grasgruber, Pavel, Pokolení Adama a Evy 1-5,
National Geography Česko,
2008
Vencl,
Slavomil – Fridrich, Jan,
Archeologie pravěkých Čech 2.
Paleolit
a mezolit, Praha 2007
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}